本研究は小脳プルキンエ細胞で長期抑圧(LTD)を誘発する非線形なCa2+濃度変化をCa2+緩衝作用の点から検討するため、Ca2+緩衝能の定量化を低親和性カルシウム蛍光指示薬と高親和性ケイジドカルシウム化合物を用いカルシウム画像解析を行うことで計ったものであり、下記の結果を得ている。 1.培養小脳プルキンエ細胞を-70mVに膜電位固定しておき、0.5秒間0mvに脱分極し、低親和性カルシウム蛍光指示薬BTCを用いて細胞内Ca2+濃度の上昇を定量した。すると約1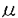 Mを閾値としてシグモイダル、即ち2相に変化することがわかった。また一旦上昇したCa2+濃度が減衰する際にも同様に2相に経過することがわかった。このCa2+濃度上昇は膜電位依存性カルシウムチャンネル阻害剤AgalVaを用いることでほぼ完全に阻害された。また、同様な非線形的なカルシウム応答はプルキンエ細胞の登上線維入力を模倣する低頻度(1-4Hz)の短い脱分極パルス(10-50ms)の繰り返し刺激でも見られた。これらの非線形的なカルシウム応答は高親和性カルシウム蛍光指示薬Fura-2を用いても観察されるが、Fura-2のCa2+による飽和のために非線形応答の定量的測定は困難であることがわかった。 Mを閾値としてシグモイダル、即ち2相に変化することがわかった。また一旦上昇したCa2+濃度が減衰する際にも同様に2相に経過することがわかった。このCa2+濃度上昇は膜電位依存性カルシウムチャンネル阻害剤AgalVaを用いることでほぼ完全に阻害された。また、同様な非線形的なカルシウム応答はプルキンエ細胞の登上線維入力を模倣する低頻度(1-4Hz)の短い脱分極パルス(10-50ms)の繰り返し刺激でも見られた。これらの非線形的なカルシウム応答は高親和性カルシウム蛍光指示薬Fura-2を用いても観察されるが、Fura-2のCa2+による飽和のために非線形応答の定量的測定は困難であることがわかった。 2.繰り返し脱分極刺激の際にカルシウム電流の大きさには変化がないのに対し、 [Ca2+]iは途中から大きく変化したため、非線形的なカルシウム応答は流入するカルシウム電流が増えるためではないことがわかった。 [Ca2+]iは途中から大きく変化したため、非線形的なカルシウム応答は流入するカルシウム電流が増えるためではないことがわかった。 3.細胞内ストアからのCa2+放出効果を除くため、細胞内ストアへのSERCA(Sarco-endoplasmic reticulum Ca2+)pumpの阻害剤であるCyclopiazonic acid(CPA)を還流液に加えにも拘わらず繰り返し脱分極刺激には大きな影響を受けることなくやはり非線形的に大きくCa2+濃度は上昇することがわかった。さらにRyRの阻害剤であるRuthenium redもパッチピペット内に加えておいてもカルシウム応答は変わらず、これは既に報告されているスライス標本の結果とは異なるため、非線形的なカルシウム応答には細胞内ストアからのCa2+放出はほとんど関係ないことがわかった。 4.Ca2+で飽和した高親和性ケイジドカルシウム化合物DMNPE-4を直接プルキンエ細胞に導入し、6%の破壊率で繰り返し紫外光を照射して一定量ずつのCa2+を細胞内に放出させてみたところ、繰り返し脱分極刺激のときと類似した非線形的Ca2+濃度の上昇が見られた。 5.上記で得られたデータを定量的に解析した結果、プルキンエ細胞には高親和性と低親和性の2種類のCa2+緩衝能が存在することがわかった。高親和性緩衝能(KCa=0.37M)はHill係数が2で現される協調的なCa2+結合部位をもち、高濃度(0.36mM)存在し、これが飽和することでCa2+濃度上昇の非線形性が起こると結論された。このケイジド試薬を用いた方法論の有効性は、高親和性Ca2+結合蛋白質の発現していない牛副腎髄質細胞において検証された。またこの高親和性緩衝能はこの細胞に多量に発現しているCa2+結合蛋白質であるcalbindin-D28kと生化学的特性が類似することもわかった。 6.細胞体において脱分極刺激直後のCa2+濃度の緩和過程を定量的に検討し、細胞内に内在する2種類のCa2+結合部位の拡散係数を求めた。その結果、細胞内Ca2+は濃度が1M以下の時はそれ以上の時に較べて圧倒的に拡散しやすいことがわかり、拡散係数は高親和性Ca2+結合部位が約80m2/s,低親和性のそれは15m2/s以下であることがわかった。 7.高親和性緩衝能が存在することによりCa2+濃度が1M以下の時にはCa2+濃度変化の時定数が長くなり、その緩衝部位が拡散することによりCa2+の見かけの拡散定数は大きくなることがわかった。即ち登上線維入力のような低頻度のシナプス入力刺激でも十分にCa2+が蓄積してプルキンエ細胞の機能調節には有効に働けることがわかり、Ca2+は例えばcAMP同様の拡散性のセカンドメッセンジャーとして働けることがわかった。 8.高親和性Ca2+緩衝能が飽和するとカルシウムシグナルの非線形性が生じるが、さらに高親和性結合部位が拡散することを考慮に入れるとその効果がより強くなるだけでなくより狭い時間幅でのみ起きることがわかった。そしてCa2+濃度が1M以上になると固定した低親和性Ca2+結合能が寄与し、空間的に限局した部位でカルシウム応答が生じるようになるため、プルキンエ細胞における長期抑圧(LTD)を誘発する時のようなシナプスにおける複数のカルシウムシグナルの同期検知が可能であることがわかった。 以上、本論文は小脳プルキンエ細胞のカルシウム画像解析から内在するCa2+緩衝能の定量化を試みたものであり、高親和性で且つ拡散性と低親和性で且つ固定性の2種類のCa2+結合分子が存在することを明らかにした。本研究はこれまで未知に等しかった神経細胞に内在するCa2+結合分子の役割を明らかにした点で、今後Hebb型学習の際に生じるシナプス同時入力の検出機構などの脳内計算過程の解明に重要な貢献をなすと考えられ、学位の授与に値するものと考えられる。 |