中枢神経系の様々な神経伝達物質によるシナプス前抑制機構の細胞メカニズムとして、シナプス前末端カルシウム・チャネルの抑制、シナプス前末端カリウム・チャネルの活性化、シナプス前末端のカルシウム流入以降の開口放出機構の抑制が提案されている。最近、代謝型グルタミン酸受容体或いはGABAB受容体を介するシナプス前抑制が専らシナプス前末端のカルシウム電流の抑制によって生じることが明らかになった。これらの代謝型受容体を介するシナプス前抑制は、多くのシナプスにおいて百日咳毒素感受性Gi/oクラスの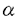 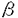 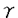 サブユニットからなる3量体型Gタンパクによって媒介されていると考えられている。しかし、シナプス前抑制にGタンパクが関与していることの直接的証拠は示されていない。 サブユニットからなる3量体型Gタンパクによって媒介されていると考えられている。しかし、シナプス前抑制にGタンパクが関与していることの直接的証拠は示されていない。 シナプス前末端におけるシナプス前抑制現象のメカニズムを実験的に明らかにするには、大きなシナプス前末端から直視下に記録を行なうことが望まれる。脳幹の台形体内側核(MNTB)にはthe calyx of Heldと呼ばれる非常に大きなシナプス前終末が存在する。本研究ではこの標本を用いてシナプス前抑制機構の解明を試みた。グアニン3リン酸類似体もしくは3量体型Gタンパクのサブユニットをシナプス前末端the calyx of Heldに注入して、GABAB受容体を介するシナプス前抑制機構を検討したところ、Gタンパクサブユニットがシナプス前抑制を媒介することを示唆する結果が得られた。 厚さ250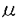 mのラット脳幹の横断面スライスを作製し、双極電極を用いてMNTBへのシナプス入力繊維を0.1Hzで細胞外刺激し、このときに誘発される興奮性シナプス後電流(EPSC)をホールセル電位固定法でMNTBの細胞から記録した。GABAB受容体アゴニストbaclofen(20M)を潅流投与するとEPSCの振幅は可逆的に減少した(72.5±4.3%;n=8)。また、厚さ150mの脳幹横断面スライスを作製し、シナプス前末端と後細胞の両方から同時にホールセル記録を行い、シナプス前末端の活動電位によって誘発されるEPSCを測定した。同時ホールセル記録によるEPSCはbaclofenの投与によって同程度に減少した(78.5±0.71%;n=4)。 mのラット脳幹の横断面スライスを作製し、双極電極を用いてMNTBへのシナプス入力繊維を0.1Hzで細胞外刺激し、このときに誘発される興奮性シナプス後電流(EPSC)をホールセル電位固定法でMNTBの細胞から記録した。GABAB受容体アゴニストbaclofen(20M)を潅流投与するとEPSCの振幅は可逆的に減少した(72.5±4.3%;n=8)。また、厚さ150mの脳幹横断面スライスを作製し、シナプス前末端と後細胞の両方から同時にホールセル記録を行い、シナプス前末端の活動電位によって誘発されるEPSCを測定した。同時ホールセル記録によるEPSCはbaclofenの投与によって同程度に減少した(78.5±0.71%;n=4)。 この抑制にGタンパクが関与しているかどうかを調べる為に、Gタンパクの活性化阻害剤GDPS(3mM)を、ホールセル・ピペットから前末端内に注入した。GDPS存在下においてはbaclofen(20M)によるEPSCの抑制は認められなかった。引き続いて、GTPを含むピペットを用いて同一の前末端から同時記録を行ったところ、baclofenによるEPSCの抑制効果が観察された。したがって、baclofenによるシナプス前抑制には前末端のGタンパクが関与すると結論される。 次に、前末端カルシウム電流に対するbaclofenの作用を調べた。前末端をホールセル記録で-80mVに電位固定し、電位依存性ナトリウム電流及びカリウム電流を薬理学的にブロックした状態で、脱分極電位パルスによって誘発される前末端カルシウム電流を記録した。前末端カルシウム電流はbaclofen(20M)の潅流投与によって振幅が減少し、電流の活性化が遅くなった。 次に、ケージドGTPS(38M)を前末端に注入して、前末端カルシウム電流を記録した。このケージド化合物は紫外光の照射によって光分解し、Gタンパクを非可逆的に活性化するGTPSを放出する。したがって紫外光照射によるGタンパク活性化の前後でのbaclofenの効果を、一つの細胞で比較することができる。この状態で紫外光照射前にbaclofen(20M)を潅流投与すると前末端カルシウム電流は可逆的に減少した。紫外光を照射すると、前末端カルシウム電流の振幅が減少し、活性化が遅くなった。この状態でbaclofen(20M)を潅流投与しても前末端カルシウム電流は変化しなかった。この結果から、baclofenによる前末端カルシウム電流の抑制もGタンパクの活性化によって起こると考えられる。 一般に、代謝型受容体とその標的タンパクの間を媒介するGタンパクは、3量体型Gタンパクであると考えられている。特にGABAB受容体を介するN型カルシウム電流の抑制には、Gi/oクラスの3量体型Gタンパクが関与していることが多くの研究で示唆されている。Gi/oクラスの3量体型Gタンパクは、百日咳毒素(PTX)によってADPリボシル化され、これによってGi/oクラスの活性化が特異的に阻害される。まず、Gi/oクラスの3量体型Gタンパクが、baclofenによるシナプス前抑制又は前末端カルシウム電流の抑制に関わっているかどうかを、PTXを用いて調べた。厚さ350Mの脳幹スライスを5-10mg/mlPTX存在下或いは非存在下、37.5℃で7時間以上保存した後に、MNTBの細胞から細胞外刺激で誘発されるEPSCを記録した。PTX処理したスライスにけるEPSCの抑制の程度(39.8±9.0%;n=11)は、コントロール(71.8±2.6%;n=7)と比較して有意に小さかった。N-エチルマレイミド(NEM)は、PTXがADPリボシル化するのと同じ部位でGi/oクラスの3量体型Gタンパクをアルキル化し、これによってGi/oクラスの活性化を特異的に阻害する。NEM(1mM)を前末端に注入して前末端カルシウム電流を記録し、baclofenによる抑制に対する影響を調べた。NEM存在下では、baclofenによる前末端カルシウム電流の抑制は生じなかった。これらのPTX及びNEMの結果は、baclofenによるシナプス前抑制にGi/oクラスの3量体型Gタンパクが関与することを示唆する。 次に、GBを前末端に注入しカルシウム電流への影響を検討した。Gの注入によって、前末端カルシウム電流はbaclofen投与時と同様に徐々に減少し、電流の活性化も遅くなった。煮沸により不活性化したGは前末端カルシウム電流に影響しなかった。 もしもGの注入によるカルシウム電流の減少が、baclofenによる前末端カルシウム電流の抑制と同じ機構で生じているとすれば、両者の効果が互いに閉塞(occlusion)されることが期待される。baclofenによる抑制効果を、ホールセル記録の開始直後とその15分後で比較した。G(200nM)の存在下では、baclofen(2M)の初回投与による前末端カルシウム電流の抑制は43.1±2.3%(n=8)であったが、その15分後の投与による抑制効果は28.9±2.7%(n=8)と、有意に減少した。したがってGとbaclofenは共通の機構を介してカルシウム電流を抑制することが示唆される。 Gタンパクによるカルシウム・チャネルの抑制は、大きな脱分極パルスによって解除され、カルシウム電流の活性化挙動が早くなり振幅が大きくなるunblockの現象が知られている。この現象が、G存在下による前末端カルシウム電流の抑制にも見られるかどうかを検討した。膜電位-80mVから様々な膜電位へ脱分極するテスト・パルスによって前末端カルシウム電流を誘発した。脱分極パルス開始の20ミリ秒前に10ミリ秒間+100mVの強い脱分極パルスを与えると、G(200nM)存在下では前末端カルシウム電流が抑制から解除された。 以上の結果より、GABAB受容体を介するシナプス前抑制はGタンパクによって媒介され、Gi/oクラス3量体型Gタンパクのサブユニットがシナプス前末端カルシウム電流を抑制することによって生じることが明らかとなった。 |