CD40はTNFRスーパーファミリーに属し、B細胞、B前駆細胞、マクロファージ、濾胞性樹状細胞、胸腺上皮細胞などの細胞表面に発現する糖蛋白質である。一方で、そのリガンドCD40Lは主に活性化T細胞に発現する。このCD40-CD40L相互作用は免疫グロブリンクラススイッチ、胚中心形成、メモリーB細胞形成などに必要であり、さらに抗原提示細胞上のCD40シグナルが細胞傷害性T細胞の活性化を誘導することが知られている。 CD40の細胞質領域(cyt)はマウスCD40(mCD40)とヒトCD40(hCD40)の間で完全に保存されているC末端側領域a.a.246-269(cyt-C)とホモロジーの低いN末端側領域a.a.216-245(cyt-N)とに分けることができる。CD40による多様な免疫反応の一部はNF-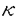 Bが担っていると考えられているが、我々はCD40Lと共にcyt-N、cyt-Cそれぞれを過剰発現させるとNF-B活性化がみられることを以前示している。CD40には酵素活性をもつような特徴的領域がないが、少なくとも4種類のTRAFファミリーTRAF2、TRAF3、TRAF5、TRAF6がCD40細胞質領域に結合することを、我々及び他のグループが示してきた。これらの内、TRAF2,5,6はそれぞれの過剰発現によってNF-Bを活性化できる。CD40内でのTRAF結合部位についてはTRAF2,3,5に関して詳しく調べられており、TRAF2,3,5の結合にはcyt-Cを必要とすること、この領域のThr254をAlaに置換(T254A)するとTRAF2,3,5の結合が失われることが報告されている。しかし、TRAF6に関しては結合部位が確定されておらず、私はこの部位の決定を試みた。まず、cyt-N、cyt-CそれぞれとTRAF6との結合をin vivoでみた結果、TRAF6はcyt-Nのみに結合することを明らかにした。次に、cyt-N内のTRAF6結合部位を限定するため、cyt-Nにおいてマウスとヒトとで保存された残基をAlaに置換していった結果、Glu235をAlaに置換した変異体(E235A)でTRAF6との結合が完全に失われた。mCD40でも対応する変異E239AでTRAF6の結合が失われた。さらに、cyt-NによるNF-B活性化はE235A変異で、cyt-CによるNF-B活性化はT254A変異で完全に抑制されることを明らかにした。これらから、cyt-NによるNF-B活性化にはTRAF6の結合が、cyt-Cによる活性化にはTRAF2またはTRAF5の結合が必要であることが示唆された。NF-Bレポーターアッセイでみる限り、cyt-Nによる活性化とcyt-Cによる活性化の量的な比は約7対3であり、顕著な量的差が存在した。実際、CD40全長(cyt)にE235A変異を導入した場合のNF-B活性化は野性型の30%程度まで減少するのに対し、T254Aの導入ではわずかな減少しか認められなかった。 Bが担っていると考えられているが、我々はCD40Lと共にcyt-N、cyt-Cそれぞれを過剰発現させるとNF-B活性化がみられることを以前示している。CD40には酵素活性をもつような特徴的領域がないが、少なくとも4種類のTRAFファミリーTRAF2、TRAF3、TRAF5、TRAF6がCD40細胞質領域に結合することを、我々及び他のグループが示してきた。これらの内、TRAF2,5,6はそれぞれの過剰発現によってNF-Bを活性化できる。CD40内でのTRAF結合部位についてはTRAF2,3,5に関して詳しく調べられており、TRAF2,3,5の結合にはcyt-Cを必要とすること、この領域のThr254をAlaに置換(T254A)するとTRAF2,3,5の結合が失われることが報告されている。しかし、TRAF6に関しては結合部位が確定されておらず、私はこの部位の決定を試みた。まず、cyt-N、cyt-CそれぞれとTRAF6との結合をin vivoでみた結果、TRAF6はcyt-Nのみに結合することを明らかにした。次に、cyt-N内のTRAF6結合部位を限定するため、cyt-Nにおいてマウスとヒトとで保存された残基をAlaに置換していった結果、Glu235をAlaに置換した変異体(E235A)でTRAF6との結合が完全に失われた。mCD40でも対応する変異E239AでTRAF6の結合が失われた。さらに、cyt-NによるNF-B活性化はE235A変異で、cyt-CによるNF-B活性化はT254A変異で完全に抑制されることを明らかにした。これらから、cyt-NによるNF-B活性化にはTRAF6の結合が、cyt-Cによる活性化にはTRAF2またはTRAF5の結合が必要であることが示唆された。NF-Bレポーターアッセイでみる限り、cyt-Nによる活性化とcyt-Cによる活性化の量的な比は約7対3であり、顕著な量的差が存在した。実際、CD40全長(cyt)にE235A変異を導入した場合のNF-B活性化は野性型の30%程度まで減少するのに対し、T254Aの導入ではわずかな減少しか認められなかった。 さらに、cyt-Cとcyt-NによるNF-B活性化には量的な違いだけでなく質的な違い、すなわち、両者が異なるシグナル経路を用いている可能性を示した。NIK(NF-B-inducing kinase)はサイトカインによるNF-B活性化に必須のMAPKKKとされ、NIKのkinase negative変異体(NIK-KN)の過剰発現はNIKを介したNF-B活性化経路を遮断することが知られている。そこで、cyt-N、cyt-CによるNF-B活性化に対するNIK-KNの影響を調べたところ、NIK-KNによってcyt-CによるNF-B活性化はほぼ完全に抑制されるのに対し、cyt-Nによる活性化は部分的に抑制されるに過ぎなかった。さらに、cyt-CはTRAF2またはTRAF5を介して、cyt-NはTRAF6を介してNF-Bを活性化すると考えられることから、各TRAFの過剰発現によるNF-B活性化へのNIK-KNの影響をみたところ、TRAF2及びTRAF5による活性化はNIK-KNによりほぼ完全に抑えられるが、TRAF6による活性化は部分的に抑制されるだけであった。これらの結果から、cyt-CはTRAF2あるいはTRAF5からNIKを介してNF-Bを活性化するのに対して、cyt-NによるNF-B活性化は一部NIK以外のシグナル経路を用いている可能性が示唆された。cyt-Nとcyt-Cが異なるシグナル経路を用いていることは免疫系でのCD40の多彩な機能を反映しているのかもしれない。 自己免疫の危険を避けるため、免疫系は自己認識するB細胞を排除する機構を備えている。この排除機構を解明することは、自己免疫疾患などの今日の免疫学が抱える大きな課題を解決することにつながる。マウスB細胞株WEHI-231は、抗原受容体sIgMの架橋によって細胞死が誘導され、自己認識したB細胞の排除機構のモデルとして研究されている。sIgM架橋されたWEHI-231は、やがて細胞周期のG1期にアレストされ、その後アポトーシスが誘導される。このG1アレストとアポトーシスを抑制する活性がCD40を介したシグナルに存在することが示され、B細胞の排除と選択の機構にCD40-CD40L相互作用が深く関与することが明らかになってきた。私は、WEHI-231での細胞死と生存の制御におけるCD40シグナルの作用機構について、TRAF蛋白質の観点から明らかにしようと試みた。上述のようにTRAF6はcyt-Nに、TRAF2,3,5はcyt-Cに結合し、それぞれについて結合を消失させるアミノ酸変異も同定できたことから、まず様々なhCD40の変異体を恒常的に発現したNVEHI-231株を樹立し、それぞれについて細胞死抑制活性を調べた。その結果、cyt-N単独では細胞死抑制能をもたない(これは石田らが既に報告済)のに対し、cyt-Cは単独で細胞死抑制能をもち、しかもこの能力はT254A変異で完全に失われることが明らかとなったことから、TRAF2、TRAF3、TRAF5のいずれかがこのシグナルを伝達する分子である可能性が強く示唆された。他方、CD40全長(cyt)にE235A変異を導入したcyt/E235Aは野性型cytと全く同程度の細胞死抑制能を示したことから、TRAF6のCD40への結合は、この活性に必要ではないと考えられた。ただし、cyt-C/T254Aは細胞死を全く抑制できないが、cyt/T254Aは弱いながら抑制能をもつことから、cyt-NはTRAF6以外の何らかの分子機構を介して細胞死抑制に一部寄与していることが示唆された。s-IgM架橋で誘導されるアレスト・細胞死の抑制にはNF-B活性が必要であるという報告がいくつかなされているが、NF-B活性化の量的な程度はcyt-Cよりもcyt-Nの方が大きいことから、cyt-Nとcyt-Cによって活性化されるNF-Bには質的差異があり、cyt-Cから誘導されるタイプのNF-B活性が細胞死抑制能に必要であると推測される。 また、sIgM架橋によるG1アレスト誘導の際に細胞周期の主要な調節蛋白質であるCyclin dependent kinase 4(Cdk4)の蛋白質量が減少し、Cdkの阻害蛋白質の一つであるp27/Kip1の蛋白質量が増加するが、この変化はCD40刺激によって無刺激細胞と同レベルに回復することが知られている。私は、p27及びCdk4の発現量の調節機構にはmRNAレベルと蛋白質レベルの2つの調節が存在することを示す。 私のこれまでの研究によってCD40内でのTRAF6結合部位を決定することができた。そこで、CD40内のTRAF6結合部位周辺の配列をもとに、TRAF6結合配列に類似した配列をもつ蛋白質をデータベース上で検索した結果、NK細胞に発現するMHCクラスI受容体が候補として得られた。この受容体は、自己MHCクラスIを認識し、NK細胞が自己細胞を殺さないように抑制性シグナルを伝達するものである。そこでMHCクラスI受容体の内、NKAT2と各種TRAFとの結合を調べた結果、NKAT2とTRAF6が結合した。また、Fc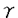 RIIb1細胞外領域とNKAT2細胞内領域をもつキメラ受容体を293T細胞に過剰発現させたところ、NF-B結合配列依存的レポーターの活性化が観測された。これらから、NKAT2はTRAF6を介してNF-Bを活性化している可能性が示唆された。 RIIb1細胞外領域とNKAT2細胞内領域をもつキメラ受容体を293T細胞に過剰発現させたところ、NF-B結合配列依存的レポーターの活性化が観測された。これらから、NKAT2はTRAF6を介してNF-Bを活性化している可能性が示唆された。 |