本研究はCCケモカインと比較して発現が困難で解析の遅れていたSDF-1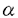 およびSDF-1 およびSDF-1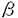 をセンダイウイルス(SeV)ベクターを用いて真核細胞で大量発現させ、その抗HIV活性を詳しく検討することを目的としたもので、下記の結果を得ている。 をセンダイウイルス(SeV)ベクターを用いて真核細胞で大量発現させ、その抗HIV活性を詳しく検討することを目的としたもので、下記の結果を得ている。 1.マウスより得られたSDF-1および遺伝子をヒト型に改変し、Katoら(Genes.Cells.1:569-579.1996)の開発したセンダイウイルスベクターに挿入し、感染性組み換えセンダイウイルス(rSeV)を作成した。CV-1細胞におけるSDF-1の転写はNorthern blotにより24時間から96時間の範囲で確認できた。上清をSDS-PAGEで解析したところ分子量約8kD付近に主要なバンドが存在し、これは作成した抗SDF-1抗体に特異的に反応した。 2.rSeVをトリ胎児繊維芽細胞に感染させ、感染72時間後の培養上清を回収した。培養上清は口径5 mのフィルターを隔てて濃度依存的に白血球を走化させた。またCXCR4をコレセプターとして使用するT細胞指向性HIV-1 NL43,SF33,TK11株に対して顕著な増殖抑制活性を示す一方、CCR5をコレセプターとして使用するHIV-1株およびSIVmac239に対しては抑制活性を示さず、CXCR4を介したHIV-1感染を特異的に阻害することが確認された。これらは報告されているSDF-1の性質と一致した。 mのフィルターを隔てて濃度依存的に白血球を走化させた。またCXCR4をコレセプターとして使用するT細胞指向性HIV-1 NL43,SF33,TK11株に対して顕著な増殖抑制活性を示す一方、CCR5をコレセプターとして使用するHIV-1株およびSIVmac239に対しては抑制活性を示さず、CXCR4を介したHIV-1感染を特異的に阻害することが確認された。これらは報告されているSDF-1の性質と一致した。 3.さらにガラクトシダーゼ発現を指標として行う細胞融合実験で、SDF-1を含む培養上清がHIV-1特異的細胞融合を阻止することも確認された。一方、HIV-1のLTR下流にルシフェラーゼを挿入したプラスミドおよびTatの発現プラスミドを細胞に同時にトランスフェクトする系ではSDF-1を含む培養上清の有無によりルシフェラーゼの発現に差異がみられなかったことより、SDF-1はHIV-1ゲノムの転写には影響を与えないことが確認された。 4.培養上清は顕著にカルシウムイオンの末梢血単核細胞内への流入をひきおこし、これはGタンパク阻害剤Pertussis Toxin(PTX)により阻止された。しかしながらPTXのみでHIV-1の増殖抑制はおこらなかったのに加え、PTXによりSDF-1の有するHIV-1増殖抑制は解除されず、Gタンパクを介したシグナル伝達系はHIV-1の細胞侵入とSDF-1によるHIV-1増殖抑制には関与していないことが示唆された。 5.培養上清中のSDF-1をヘパリンカラムを用いて精製を行った。10mlの培養上清から36gのSDF-1を精製した。培養上清と比較して比活性は3倍に上昇し、収率は9%であった。活性画分中のSDF-1はSDS-PAGE上で約8kDの単一のバンドとして移動し、このバンドは抗SDF-1抗体に特異的に反応した。N末端アミノ酸配列解析でこのバンドのN末端アミノ酸配列が報告されているSDF-1のそれと一致した。 6.SDF-1およびSDF-1両者の生物活性の相違点は未知であった。本研究では両者それぞれを上記の系で発現および精製し、白血球走化活性およびHIV-1増殖抑制活性について検討したところ両者の間に活性の相違は殆どみられなかった。 7.SeV/SDF-1の遺伝子のN末端にPCRで変異を導入した。導入した変異はKPV→APV(AP),KPR(KPR),KKKPV(KKKP),HHHHHHKPV(His6〜SDF)の4種類である。前述の方法と同様にして組み換えウイルスを作成し、変異SDF-1を発現した。白血球走化活性および抗HIV活性を検討したところ前者についてはKKKPが増強されている傾向がみられた。後者についてもKKKPおよびHis6-SDFが増強されている傾向があり、100ng/mlの処理でKKKPはSDF-1に比べて有為に増強されていることが分かった。 以上、本論文はSeVによるSDF-1およびSDF-1の発現と簡便な精製系の確立を行い、両分子の白血球走化活性およびHIV-1増殖抑制活性には差異が殆どみられないことを明らかにした。またSDF-1の改変により、SDF-1のN末端の伸長による活性の増強の可能性を示唆した。本研究は抗HIV/AIDS治療薬の新たな作用点の候補であるHIV-1の細胞侵入を防御するという目的において、考え得るいくつかの方法のひとつであるケモカインをベースとした薬剤の開発において必要となるであろう、タンパク質の新たな簡便な発現精製系を提示した点について、さらにケモカインについての詳細な情報の蓄積の必要性という面において、SDF-1のN末端の伸長による活性の増強の可能性を提案した点において重要であり、学位の授与に値するものと考えられる。 |