赤内型マラリア原虫に対する免疫防御機構としてはサイトカイン、NK細胞による細胞障害活性、マクロファージ等のよる貪食、抗体等様々な免疫応答反応の関与が指摘されている。本研究では赤内型マラリア原虫のマウスモデルとして、Plasmodium berghei(P.berghei)の強毒株NK65およびこの強毒株をX線照射により弱毒化した自然治癒株XATを用いて、その免疫防御機構の解明を試みた。近年、赤内型マラリア原虫に対する免疫防御機構にはCD4+T細胞とIFN-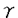 の関与が指摘されているが、それ以外にもNK細胞やNitric oxide(NO)もまた、赤内型マラリア原虫に対する感染防御に関与すると考えられている。本研究では、まず強毒株NK65および自然治癒株XAT感染における脾臓内でのIL-12、IFN-等のサイトカインの誘導の機構の解明を試み、次にIFN-の下流に位置するNK細胞の活性化とマクロファージから産生されるNOが自然治癒株XATに対する感染防御に関与するか否かの検討を行い、さらに、マクロファージの感染赤血球に対する貪食能の関与について検討した。 の関与が指摘されているが、それ以外にもNK細胞やNitric oxide(NO)もまた、赤内型マラリア原虫に対する感染防御に関与すると考えられている。本研究では、まず強毒株NK65および自然治癒株XAT感染における脾臓内でのIL-12、IFN-等のサイトカインの誘導の機構の解明を試み、次にIFN-の下流に位置するNK細胞の活性化とマクロファージから産生されるNOが自然治癒株XATに対する感染防御に関与するか否かの検討を行い、さらに、マクロファージの感染赤血球に対する貪食能の関与について検討した。 本研究で明らかになったのは以下の6点である。 1.P.berghei XAT感染は脾臓におけるIL-12産生とそれに引き続くIFN-産生を増強させることが明らかとなった。また、抗IL-12抗体、抗IFN-抗体を用いたin vioでの中和実験結果より脾臓で産生されたIL-12およびIFN-は、P.berghei XATに対して感染防御的に作用することが示された。一方、P.bergheiNK65感染も脾臓および肝臓においてIL-12産生とそれに引き続くIFN-産生を増強させるが、感染防御よりむしろ肝障害を引き起こし、病態増悪的に作用すると考えられた。 2. P. berghei XAT感染マウスの脾細胞においてはNK細胞の細胞障害活性が感染後4日をピークとして著明に増強するが、P.bergheiNK65感染マウスにおいては感染後4日に若干の増強を示したのみであった。そこで、P.bergheiXAT感染マウスに対してin vivoにおけるNK細胞除去の目的でNK1.1抗体の投与を行ったところ、NK細胞の細胞障害活性は著明に減少したが、parasitemiaの経時的な変化はコントロール抗体投与マウスと比較してもほとんど変わらず、マウスは全例治癒した。また、この時抗NK1.1抗体投与を行ったマウス脾細胞におけるIFN-の産生にはほとんど変化が見られなかった。これらの結果からP.berghei XAT感染防御におけるNK細胞の関与は低いと考えられた。 3. P.berghei XAT感染マウスの脾細胞をin vitroで培養し、その上清中のNO2を測定したところ、感染6日後をピークとしてNO2産生が認められたが、P.bergheiNK65感染マウスではNO2産生はほとんど認められながった。そこでNO産生の感染防御への関与を調べるためにiNOSのノックアウトマウスを用いてP.bergheiXATの感染実験を行ったが、iNOSのノックアウトマウスにおけるparasitemiaの経時的な変化はコントロールマウスと比較してほとんど変わらず全例治癒した。この結果から、NOのP.bergheiXAT感染防御における関与は低いと考えられた。 4.P.berghei XAT感染マウス脾細胞からのIFN-の産生は、感染後4日をピークとして認められる。しかし、CD4ノックアウトマウスを用いてP.berghei XAT感染実験を行うと脾細胞からのIFN-の産生がほとんど認められずマウスは全例死亡した。この結果からCD4+T細胞から産生されるIFN-がP.berghei XATに対する感染防御に深く関与していると考えられた。 5.P.bergheiXAT感染マウスにマクロファージの貪食阻害物質であるCGN(carrageenan)を投与したところマウスは高いparasitemiaを示し死亡した。またCGNを投与した感染マウスの脾附着細胞をin vitroでFITC蛍光ビーズとともに培養したところ、蛍光ビーズに対する貪食能がコントロールマウスに比較して著しく低下していた。これらの結果より脾臓のマクロファージの貪食能がP.berghei XATに対する感染防御に関与していると考えられた。 6.P.berghei XATに対する免疫血清で感作した感染赤血球に対するP.berghei XAT感染マウス脾臓マクロファージの貪食能を測定したところ、感染後7日をピークとして貪食能の増強が認められた。この結果より抗体依存性の貪食能がP.berghei XAT感染に対する感染防御に関与していると考え、Fcレセプターchain欠損マウスを用いて感染実験を行ったところ、高いparasitemiaを示し全例死亡した。また、Fcレセプターchain欠損マウス脾臓マクロファージの感染赤血球に対する抗体依存性貪食能の増強はほとんど認められなかった。Fcレセプターchain欠損マウスにおけるIFN-産生能、抗体産生能はコントロールマウスと比較して差が認められなかった。これらの結果より、P.berghei XATに対する感染防御にはFcレセプターが介在する抗体依存性の貪食能が関与していると考えられた。また、IFN-ノックアウトマウスもP.berghei XATに対して高い感受性を示すが、このマウスにおける感染赤血球に対する脾臓マクロファージの貪食能および血清中のIgG2aサブクラスの抗体価は低下しているため、抗体依存性貪食にはIFN-による脾臓マクロファージおよびB細胞の活性化が必要であると考えられた。 以上の結果より、弱毒株P.berghei XAT感染マウスにおいては初期に脾臓内で内在性のIL-12の誘導が起こりIFN-が誘導産生され、このIFN-がP.berghei XATに対する感染防御に重要であると考えられた。また、強毒株P. berghei NK65感染も脾臓および肝臓においてIL-12産生とそれに引き続くIFN-産生を増強させるが、これらは感染防御よりむしろ肝障害を引き起こし、病態増悪的に作用すると考えられた。 弱毒株P.berghei XAT感染マウスの脾臓においてはIFN-によるNK細胞の細胞傷害活性の増強やNO産生の誘導が起きるが、それらの宿主の感染防御への関与は少なく、抗体価の上昇とともにFcレセプターが介在する、活性化されたマクロファージによる感染赤血球に対する抗体依存性貪食の増強が感染防御に重要であることが明らかになった。また、IFN-の産生細胞としてはCD4+T細胞が最も重要であると考えられた。 |