本研究はヒトT細胞白血病細胞株MOLT-4細胞の放射線誘発アポトーシスの分子機構を解析するため、セラミド生成を触媒する酵素である酸性スフィンゴミエリナーゼの阻害剤やMOLT-4細胞由来のX線抵抗性雑種及び、細胞死抑制遺伝子bcl-2を導入した形質転換株を用いた系にて、放射線照射後の細胞死に至るメカニズムの解析を試みたものであり、下記の結果を得ている。 1.MOLT-4細胞はX線に対して線量依存的にアポトーシスを起こす。セラミド生成を触媒する酸性スフィンゴミエリナーゼの阻害剤(D609)で細胞を前処理することにより、MOLT-4細胞のアポトーシスが有意に抑制された。またMOLT-4細胞は細胞膜透過性の合成セラミド(C2-セラミド)に対して濃度依存的にアポトーシスを起こした。さらにX線照射後、MOLT-4細胞ではp53の蓄積が起こるが、C2-セラミド処理後ではp53の蓄積が見られないこと、D609処理はX線照射後のp21/WAF-1の誘導は阻害しないことが明らかになった。これらのことは、X線照射後のアポトーシス誘発機構にセラミド生成及びp53非依存性のセラミドシグナル伝達経路の活性化が必要であることが示唆された。 2.MOLT-4細胞、マウスFM3A細胞及びその両親株から作製したX線抵抗性放射線雑種(radiation-hybrid clones)、MOLT-4にbcl-2遺伝子を導入した形質転換株(MOLT/mbcl-2)を用い、X線やC2-セラミド誘発アポトーシスに対する感受性を解析したところ、radiation-hybrid clonesやMOLT/mbcl-2細胞はX線誘発アポトーシスに抵抗性を示し、さらにradiation-hybrid clonesの1つは、FM3AやMOLT/mbcl-2細胞と同様に高いセラミド抵抗性を示した。またradiation-hybrid clones、MOLT/mbcl-2の各細胞におけるX線照射後のp53の蓄積は親株であるMOLT-4細胞と同様に見られた。したがって、C2-セラミド誘発アポトーシスにも交差抵抗性を示すradiation-hybrid cloneの結果からも、X線照射誘発アポトーシスにおいてセラミド伝達経路が重要な働きをしていることが示唆された。 3.X線照射後のstress-activated protein kinase(SAPK/JNK)の活性化について調べたところ、MOLT-4細胞では刺激後、持続的で高い活性が見られた。一方、酸性スフィンゴミエリナーゼの阻害剤(D609)で前処理またはBcl-2過剰発現したMOLT-4細胞ではX線照射後のSAPK/JNKの活性化が抑制されていた。セラミド誘発アポトーシスに交差抵抗性を示すradiation-hybrid cloneでは照射後、一過性の活性化しか見られなかった。C2-セミド処理後、MOLT-4やBcl-2過剰発現したMOLT-4細胞では持続的で高い活性が見られたのに対し、セラミド誘発アポトーシスに交差抵抗性を示すradiation-hybrid cloneでは一過性の活性化のみ見られた。これらのことはBcl-2はX線誘発SAPK/JNK活性化は抑制できるが、C2-セラミド誘発SAPK/JNK活性化は抑制できない、つまりBcl-2はセラミド生成より上流でシグナル伝達経路を阻害していることが示唆された。さらに、X線、セラミド誘発アポトーシス抵抗性を示すradiation-hybrid cloneでは一過性のSAPK/JNK活性化しか見れらなかったことから、アポトーシスを起こすには持続的で高いSAPK/JNK活性化の活性化が必要であること、このクローンではX線照射後セラミド生成後のシグナル伝達経路が阻害されていることが示唆された。 以上、本論文はヒトT細胞白血病細胞株MOLT-4の放射線誘発アポトーシスの分子機構において、セラミドSAPK/JNK経路が重要な働きをしていることを示した。さらにアポトーシスには一過性ではなく、持続的で高いSAPK/JNK活性が必要であること、Bcl-2はcaspase以外にセラミド生成上流でもシグナル伝達経路を阻害しうることが初めて示唆された。本研究は放射線誘発アポトーシスの分子機構、特にセラミドシグナル伝達経路の解明を通して、細胞の放射線(X線)照射に対する応答の解明について重要な貢献をなすと考えられ、学位の授与に値するものと考えられる。 | 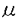 M)誘発アポトーシスに対して全く異なる感受性を示した(90.1vs.39.8 and 32.1%viability at 24 h after the treatment,for Rh-1a,MOLT-4 and Rh-5a)。Rh-1a細胞がセラミド誘発アポトーシスに対して高い交差抵抗性を示すことからもMOLT-4細胞におけるX線誘発アポトーシスではセラミド経路が重要な働きをしていることが示唆された。
M)誘発アポトーシスに対して全く異なる感受性を示した(90.1vs.39.8 and 32.1%viability at 24 h after the treatment,for Rh-1a,MOLT-4 and Rh-5a)。Rh-1a細胞がセラミド誘発アポトーシスに対して高い交差抵抗性を示すことからもMOLT-4細胞におけるX線誘発アポトーシスではセラミド経路が重要な働きをしていることが示唆された。