ヒトパピローマウイルス16型(Human papillomavirus type 16:HPV-16)に代表されるハイリスク型HPVの感染が子宮頚癌発症の最大リスクファクターである。これまでの研究で、HPV-16 E6、E7遺伝子の継続的な高発現が癌細胞の増殖能の維持と悪性形質の獲得に重要な役割を果たすことが示されてきた。HPVのE7蛋白質はpRbの機能を阻害することで、細胞のDNA合成を開始させ、細胞分裂を誘導する。一方E6蛋白質はテロメラーゼの活性化とp53の不活化により、細胞の無限増殖を可能にし、細胞DNAの損傷を固定化する。 E6、E7蛋白質の高発現がおこる機構は不明な点が多いが、細胞あたりのHPVコピー数の増大とE6、E7の転写を抑制するHPV E2遺伝子の発現阻害が原因の1つとしてあげられている。E6、E7遺伝子はlong control region(LCR)とよばれる転写調節領域によって制御されており、このLCRには多数の転写因子の結合配列が集中して存在する(図)。その中で、クラスターを形成する5力所のYYl結合配列を実験的に破壊することでE6、E7遺伝子の転写が亢進することが示されている。本研究では、実際の癌化の過程でLCRの変異によりウイルス癌蛋白質の発現が酵亢進したものがあるか調べるため、HPV-16陽性の子宮頚癌由来細胞株及び子宮頚癌生検材料を対象に、LCRのエンハンサー・プロモータ活性を詳細に検討した。 子宮頚癌由来細胞株は、細胞に組み込まれたHPVのコピー数が少ないにもかかわらず、E6、E7遺伝子が高発現しているとされるSiHa細胞と多数のHPVコピーが組み込まれたCaSki細胞を選択した。細胞あたりのHPV-16のコピー数を正確に知るため、SiHa、CaSkiのHPV-16 DNA量を定量性に優れたリアルタイムPCRシステムにより定量し、さらにフローサイトメトリーで調べた1細胞あたりのDNA含量で補正したところ、SiHaは細胞あたりHPV-16ゲノムを1〜2コピー、CaSkiはSiHaの200倍含むと推定した。また、リアルタイムPCRシステムを用いてE7 mRNAを、ウエスタンブロット法にてE7蛋白質をそれぞれ測定すると、CaSkiでは、E7 mRNA、E7蛋白質ともSiHaの約10倍発現していた。CaSkiはSiHaよりも細胞あたりのE7遺伝子の発現量は多いが、細胞に組み込まれたHPV-16ゲノム1コピーあたりの発現量はSiHaの方が高く、2つの細胞に含まれるHPVゲノムの全てから転写が行われていると仮定すると、HPVゲノムあたりの発現量はSiHaの方がCaSkiよりも20倍高いと考えられる。 SiHaにおけるE7遺伝子の高発現は、LCRの転写活性の増大が原因と予想された。そこで、HPV-16プロトタイプのLCRとSiHa、CaSkiからPCRで増幅したLCR(SiHa-LCR、CaSki-LCR)をもち、LCRの構造を厳密に保存したルシフェラーゼ発現ベクターを作製し、その転写活性を測定するとSiHa-LCRのエンハンサー・プロモータ活性は、プロトタイプのLCRやCaSki-LCRの約4倍高いことがわかった。SiHa-LCR、CaSki-LCRの塩基配列を調べたところ、SiHa-LCRにはYYl認識配列#4と#5の一部を含む38塩基対の欠失があり(図)、CaSki-LCRにはHPV-16の塩基番号12に置換変異があった。SiHa-LCRのエンハンサー・プロモータ活性の上昇は、少なくとも一部ではYYl結合領域を含む38塩基対の欠失が原因と考えられる. 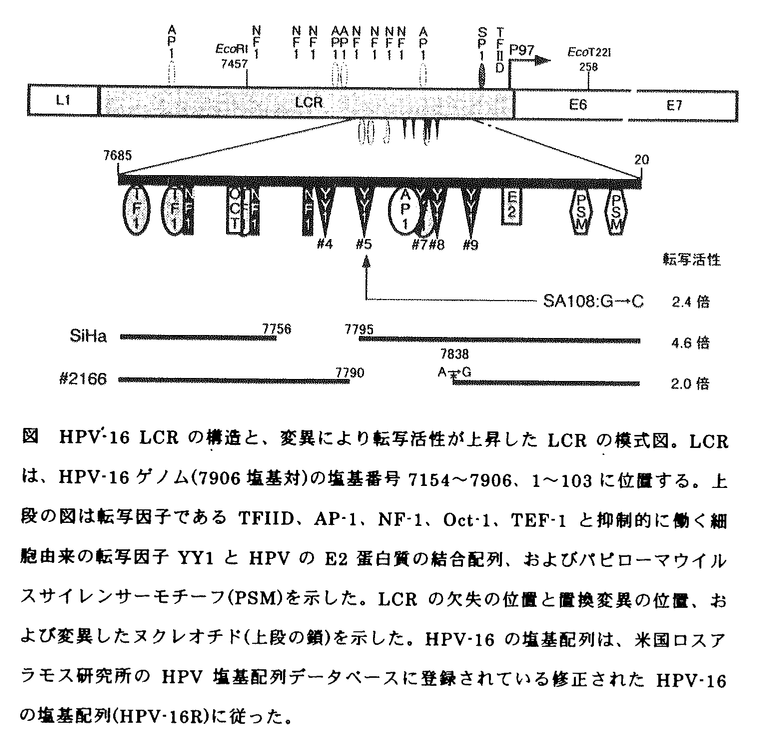 図 HPV-16 LCRの構造と、変異により転写活性が上昇したLCRの模式図。LCRは、HPV-16ゲノム(7906塩基対)の塩基番号7154〜7906、1〜103に位置する。上段の図は転写因子であるTFIID、AP-1、NF-1、Oct-1、TEF-1と抑制的に働く細胞由来の転写因子YY1とHPVのE2蛋白質の結合配列、およびパピローマウイルスサイレンサーモチーフ(PSM)を示した。LCRの欠失の位置と置換変異の位置、および変異したヌクレオチド(上段の鎖)を示した。HPV-16の塩基配列は、米国ロスアラモス研究所のHPV塩基配列データベースに登録されている修正されたHPV-16の塩基配列(HPV-16R)に従った。 図 HPV-16 LCRの構造と、変異により転写活性が上昇したLCRの模式図。LCRは、HPV-16ゲノム(7906塩基対)の塩基番号7154〜7906、1〜103に位置する。上段の図は転写因子であるTFIID、AP-1、NF-1、Oct-1、TEF-1と抑制的に働く細胞由来の転写因子YY1とHPVのE2蛋白質の結合配列、およびパピローマウイルスサイレンサーモチーフ(PSM)を示した。LCRの欠失の位置と置換変異の位置、および変異したヌクレオチド(上段の鎖)を示した。HPV-16の塩基配列は、米国ロスアラモス研究所のHPV塩基配列データベースに登録されている修正されたHPV-16の塩基配列(HPV-16R)に従った。 SiHa-LCRの転写亢進に重要な領域をさらに限定するために38塩基対よりも短い欠失を持つLCRを作成し、その転写活性を測定した。YYl結合配列に欠失のないLCRでは転写活性の増大は見られなかったが、YYl結合配列#4と#5の一部を欠失したLCRの転写活性はプロトタイプの2.5〜2.9倍高値を示し、YYl認識配列#4と#5の一部の欠失がエンハンサー・プロモータ活性を増大させていることが示された。 次に、HPV-16陽性の子宮頚癌生検材料からPCRでLCRを増幅し、その塩基配列を調べ、さらに一部はクローニングをしてエンハンサー・プロモータ活性を測定した。癌生検材料51例中、LCR内のYYl結合配列を含む領域に変異のあるものは6例あり、そのうち転写活性が増大しているものは2例あった。1例(SA108)は、YYl認識配列#5に置換変異があり、もう1例(#2166)は、YYl認識配列#5、#7、#8が欠失していた。LCRの変異によりE6、E7遺伝子の発現が亢進する症例が生検材料中にも確認され、また活性の強いプロモータからHPV E7遺伝子を発現させると、形質転換能が増強することが実験的にも示されており、実際の癌化の過程でLCRの変異により、E6、E7遺伝子の発現が亢進するものがあることが示唆された。 最近の研究でHPV-16はいくつかのサブタイプが存在することが示されており、癌生検材料51例から増幅したLCRの塩基配列をもとに、HPV-16 DNAを分類すると、ヨーロッパ型が14例(27.5%)、アジア型が35例(68.6%)、アジア・アメリカ型が2例(3.9%)であり、我が国では、HPV-16アジア型サブタイプが多数を占めることも初めて明らかになった。日本におけるアジア型の頻度は、東南アジアの頻度(27.5%)よりもはるかに高く、世界的に見てもアジア型が非常に多かった。各サブタイプのLCRの活性を比較すると、アジア・アメリカ型とアジア型のLCRの転写活性は、ヨーロッパ型の転写活性の約2〜2.5倍高かった。 子宮頚癌生検材料で細胞あたりのHPV-16コピー数を、リアルタイムPCRシステムを用いて調べ、各サブタイプ間で比較すると、ヨーロッパ型が検出される子宮頚癌の細胞あたりのHPVコピー数は、アジア型、アジア・アメリカ型が検出される癌よりも多い傾向があった。アジア型やアジア・アメリカ型よりも転写活性が低いヨーロッパ型のHPV-16に起因する子宮頚癌は、細胞あたりのHPVコピー数が増大し、E6、E7遺伝子を高発現するようになった細胞が生残した可能性が考えられる。 ヒトパピローマウイルスE6、E7癌蛋白質の発現亢進は、子宮頚癌の発生の過程で細胞が悪性形質を獲得し、それを維持するのに必要と考えられている。本研究により、実際の癌化の過程でも実験系で示されたようにLCRの変異によるE6、E7癌蛋白質の発現亢進が起こりうることが示されたが、同時に頻度が低いことから今後さらにほかの原因を探る必要がある。 |