アルツハイマー病(Alzheimer’s diseasc:AD)は初老から老年期にかけて発症する進行性の痴呆である。AD脳の病理学的特徴としては大脳皮質全体にわたる老人斑、神経原線維変化の出現と神経細胞の脱落が挙げられる。神経原線維変化はAD以外の神経疾患でも認められるが、老人斑の形成はAD脳に特異的である。 AD患者の大部分は明瞭な家族歴がないが、家族歴を持つAD患者も10%程度存在する。常染色体優性遺伝する早期発症の家族性アルツハイマー病(familial Alzheimer’s disease;FAD)の原因遺伝子として、プレセニリン(presenilin;PS)1と2が同定された。PS1遺伝子は第14染色体に、PS2遺伝子は第1染色体に位置している。この2つの遺伝子は高い相同性を持ち、膜を8回貫通する蛋白質をコードしている。2つの蛋白質は主に小胞体に局在するが、ゴルジ体にも一部分布する。しかし、その生理的な機能は不明である。PS1では50以上の点突然変異が同定されているが、PS2では2つの点突然変異が同定されている。ADを発症するVolga German家系ではPS2の141番目のアミノ酸残基のアスパラギンがイソロイシンに変異している(N141I変異)。イタリアのFAD家系ではPS2の239番目のアミノ酸残基のメチオニンがバリンに変異している。PS2の変異によるAD発症の機序は未だ不明であるが、PS1とPS2の変異はアミロイド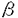 蛋白質(amyloid -protein;A)の産生に影響を与えることが知られている。A42は通常A40に比較すると低濃度分泌されているが、変異PS1とPS2を過剰発現させた培養細胞およびトランスジェニックマウス脳でA42レベルが増加することが報告されている。 蛋白質(amyloid -protein;A)の産生に影響を与えることが知られている。A42は通常A40に比較すると低濃度分泌されているが、変異PS1とPS2を過剰発現させた培養細胞およびトランスジェニックマウス脳でA42レベルが増加することが報告されている。 N141I変異を有する患者は必ず発症することから、この変異遺伝子を有するトランスジェニックマウスを確立できれば、この動物はAD発症を研究するうえでの有効なモデルになることが期待される。本研究では、野生型およびN141I変異を有するPS2cDNAを過剰発現するトランスジェニックマウスをそれぞれ作製し、PS2の脳内での動態、およびN141I変異のA産生への影響を調べた。 1.ヒト野生型と変異型PS2を発現するトランスジェニックマウスラインの確立 脳におけるヒトPS2の発現と代謝を調べるために、野生型と変異型PS2cDNAを発現するトランスジェニックマウスのラインを確立することを試みた。PS2mRNAは脳をはじめさまざまな組織で発現することが知られているので、PS2cDNAをニワトリ-アクチンプロモーターの下流に結合したPS2トランスジーンを作製した。トランスジーンを受精卵に注入し、PCRでファウンダーマウスを同定した。このようにして、野生型(W1、W2)、変異型(M1、M2)PS2cDNAをそれぞれ過剰発現するトランスジェニックマウスをそれぞれ2ラインづつ確立することが出来た。次にトランスジェニックマウスラインそれぞれの脳で発現しているヒトPS2mRNAレベルを定量RT-PCR法で調べた。トランスジェニックマウスではノントランスジェニックマウス(Non-Tg)に比べて、PS2mRNAレベルがいずれのラインでも明らかに増加していた。Non-Tgの内在性PS2mRNAを標準に比較すると、トランスジェニックマウスでは5-15倍過剰発現していた。そのなかで、W2とM1ラインはほぼ同じレベルのPS2mRNAを発現していた。 2.ヒトPS2はトランスジェニックマウス脳でNTF、CTFに切断される 脳におけるPS2とその代謝を5種類の領域特異抗体を用いたウェスタンブロット法で解析した。N末を認識する抗体2972は予測分子量50-55kDaの全長PS2の存在を示すシグナルを与えなかった。その代わりに、トランスジェニックマウスで34kDaの明瞭なバンドが認められた。この34kDaのバンドは、トランスジェニックマウスにのみ認められたことからトランスジーン由来のバンドで、全長PS2のプロテアーゼ分解によって生じたN末端断片(N-terminal fragment;NTF)と考えられる。 次にPS2のC末端領域を認識する抗体を用いて同様に分析した。PS2のループドメインに特異的な抗体では、全長PS2の存在が予想される領域に特異的なバンドを与えなかったが、Non-Tg、トランスジェニックマウス脳で22kDa断片を認識した。この断片はNon-Tgに比べ、トランスジェニックマウスでは高いレベルで存在していた。これらの結果は22kDa断片がPS2のC末端領域に由来する断片(CTF)であることを示唆する。 以上PS2の抗体を用いた結果より、トランスジェニックマウス脳で合成されたPS2はプロテアーゼの分解を受け、比較的安定な二つの断片、34kDaのNTFと22kDaのCTF、になり存在することが分かった。NTFとCTFのバンドを定量すると、W2とM1ラインでNTF、CTFレベルに差は見られなかった。 3.トランスジェニックマウス脳のTS可溶性画分のA40、A42レベル 次に、PS2mRNAの発現レベルが近いW2とM1ライン、およびNon-Tgを用いてPS2変異のA産生への影響を調べた。2、5、8カ月齢のマウス脳よりTS可溶性画分を調製し、A分子の中央部を共通に認識する抗体を捕捉抗体とし、捕捉したAをHRP標識したC末端を特異的に識別する抗体で検出するenzyme-linked immunosorbent assay(ELISA)でA40、A42それぞれのレベルを定量した。A40、A42レベルはマウス脳で変動したが各々のラインではほぼ一定の値を示した。 PS2cDNAを過剰発現すると、傾向として、Aレベルは低下した。特徴的なことはN141I変異は脳でA42レベルを上昇させることであった。特に野生型におけるA42レベルは5カ月齢以降わずかに増加するのに対し、変異型におけるA42レベルは加齢にしたがって顕著に増加した。これらのことからヒト変異型PS2トランスジェニックマウス脳では加齢にともなってA42量が増大することが明らかとなった。 4.PS2トランスジェニックマウス脳のTS不溶性画分のA40とA42レベル マウス脳より得たTS不溶性の画分をグアニジン塩酸により抽出し、ELISAでA40、A42のレベルをそれぞれ定量した。野生型マウスのTS不溶性画分のA42量はノントランスジェニックマウスと同じレベルであったが、A40よりも低い(約5倍)レベルであった。変異型マウス脳のTS不溶性画分のA42レベルは他の2ラインに比較して各月齢で有意に増加していた。 5.変異型PS2トランスジェニックマウスの低密度膜画分(detergent-insoluble,low-density membrane(LDM)fraction)でのA42レベルの増加 次にこの画分でのA42の増加がどのような細胞内コンパートメントに由来するのかを調べる目的で、TritonX-100による脳抽出液をシヨ糖密度勾配遠心法により分画し、Aの分布を調べた。この分画法により得られる低密度膜画分は他の組織ではカベオラ(caveolae)と呼ばれている部位で、コレステロールとスフィンゴリピッドの割合が高く、G蛋白質など多種類の情報伝達系に関与する分子が存在することから、情報伝達の場や細胞内輸送に関する機能が指摘されている。2カ月および8カ月齢でのA40は3ラインでほぼ同じレベルであった。しかし、15カ月齢の変異型マウスのLDM画分のA40レベルは他の2ラインに比べて有意に減少していた。一方、変異型マウス脳のLDM画分のA42レベルは野生型とノントランスジェニックマウスに比べて各年齢で有意に上昇していた。この結果、N141I変異の低密度膜ドメインでA42が上昇することが明らかとなった。 6.LDM画分の脂質解析 最後にマウス脳におけるLDM画分の脂質組成を調べた。マウスでのヒト野生型PS2の過剰発現により、LDM画分のコレステロールの量が増加し、変異型PS2を過剰発現させると、野生型PS2に比べてスフィンゴミエリン、総リン脂質、総コレステロールのレベルが有意に減少していた。 結論 1.ヒトPS2はトランスジェニックマウス脳では34kDaのNTFおよび22kDaのCTFに分解されるが、それらのレベルには各月齢の野生型および変異型PS2トランスジェニックマウス間で変化は見られない。 2.N141I変異により、脳内TS可溶性画分のA42レベルが上昇する。 3.2から8カ月齢の間に変異型トランスジェニックマウス脳のTS可溶性のA42レベルが急激に上昇するのに対して、野生型では5カ月から8カ月の間にわずかに上昇する。 4.変異型PS2トランスジェニックマウス脳の不溶性画分のA42レベルはノントランスジェニックおよび野生型トランスジェニックマウスに比べて各月齢で約2〜3倍に有意に上昇する。 5.変異型PS2トランスジェニックマウス脳の低密度膜画分のA42レベルはノントランスジェニックおよび野生型トランスジェニックマウスに比べて各月齢で有意に増加する。 6.変異型PS2トランスジェニックマウス脳の低密度膜画分のスフィンゴミエリンのレベルが野生型トランスジェニックマウスに比べて有意に減少する。 |