シナプトタグミンは膜貫通ドメインを一カ所持ち、細胞質側にPKCのC2調節領域とホモロジーを有するドメインを2つ(C2A,C2B)持つ一群のタンパク質で、現在マウス及びラットで11種類のアイソフォームの存在が知られている。最も解析されているシナプトタグミンIはシナプス小胞膜に局在し、Ca2+依存性の神経伝達物質の放出に必須であることが知られている。特にそのC2AドメインはCa2+依存的にリン脂質と結合する事が生化学的に示されており、シナプス小胞のシナプス前膜への融合過程を直接調節する事が明らかになっている(図1). 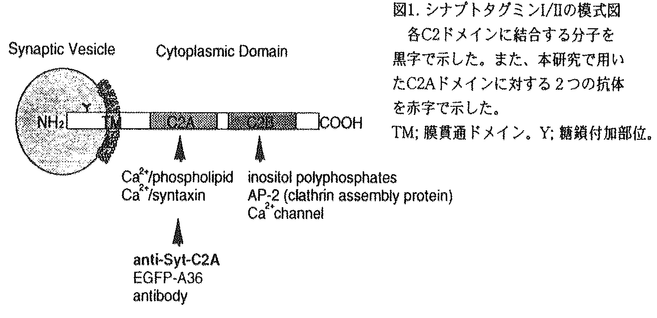 図1. シナプトタグミンI/IIの模式図各C2ドメインに結合する分子を黒字で示した。また、本研究で用いたC2Aドメインに対する2つの抗体を赤字で示した。TM;膜貫通ドメイン。Y;糖鎖付加部位。 図1. シナプトタグミンI/IIの模式図各C2ドメインに結合する分子を黒字で示した。また、本研究で用いたC2Aドメインに対する2つの抗体を赤字で示した。TM;膜貫通ドメイン。Y;糖鎖付加部位。 神経突起の先端に存在する成長円錐は神経突起伸長や軸索ガイダンスを制御し、シナプス前終末の前駆体であると考えられている。成長円錐には成長円錐小胞が豊富に存在し、突起伸長に伴う成長円錐の形質膜面積の増大の供給源と考えられている(図2)。 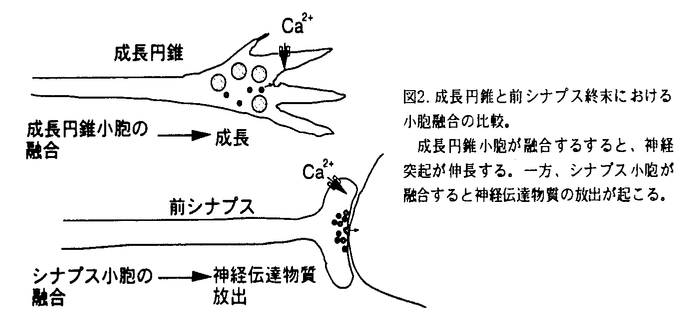 図2. 成長円錐と前シナプス終末における小胞融合の比較。成長円錐小胞が融合するすると、神経突起が伸長する。一方、シナプス小胞が融合すると神経伝達物質の放出が起こる。 図2. 成長円錐と前シナプス終末における小胞融合の比較。成長円錐小胞が融合するすると、神経突起が伸長する。一方、シナプス小胞が融合すると神経伝達物質の放出が起こる。 最近、シナプス前終末に局在し神経伝達物質放出に重要とされているSNAP25やシンタキシンがシナプス形成前の成長円錐にも局在し、突起伸長にも関与することが特異的神経毒素等を用いて明らかにされている。また成長円錐における適切なCa2+濃度が神経突起伸長には不可欠であること、神経伝達物質放出の際のCa2+センサーと考えられる。シナプトタグミンI分子が成長円錐小胞にも局在することから、私はシナプトタグミンI分子が神経突起伸長にも関与するのではないかと考えた。しかしながら、シナプトタグミンIにおいてはSNAP25やシンタキシン1Aの様に特異的神経毒素が存在しないためその検証はこれまで困難であった。そこで本論文では以下に述べる2つの独立した方法を用いて、この仮説を検証した。まず最初にマウス・シナプトタグミンI又はII(I/II)のC2Aドメインに結合し、神経伝達物質の放出を阻害する特異的機能阻害抗体(anti-STI/II-C2A)を用いて、シナプトタグミンI/IIが神経突起伸長に関与するかを検討した。次に、マウス・シナプトタグミンI/IIのC2Aドメインに対する抗体遺伝子を単離し、その遺伝子(EGFP-A36)をマウス神経細胞に導入し、突起伸長への影響を調べた。 1,神経突起伸長を調べる系として、ニワトリ11日胚の背根神経節(DRG)神経細胞を用いた。まず最初にニワトリ11日胚のDRG神経細胞の全タンパクに対してウエスタンブロット解析を行い、抗体(anti-STI/II-C2A)がニワトリ・シナプトタグミンにも特異的に交差することを確認した。また免疫染色により、ニワトリのシナプトタグミンI/IIが成長円錐に豊富に存在することを明らかにした。次に抗体をトリチュレーション法(ピペッティングによる機械的な障害で抗体を細胞内へ取り込ませる方法)で初代培養細胞に導入し、神経突起伸長への影響を調べた。その結果、抗体(二価)は神経突起の数には影響を与えず、神経突起伸長を著しく阻害した(図3)。一方、免疫前の血清から精製した対照抗体は神経突起の数、神経突起伸長のいずれにも影響を与えなかった。またFab(一価)抗体でも同様な阻害効果が得られたことから、これらの結果は二価抗体が複合体を形成することにより引き起こされたものではないと結論できた。 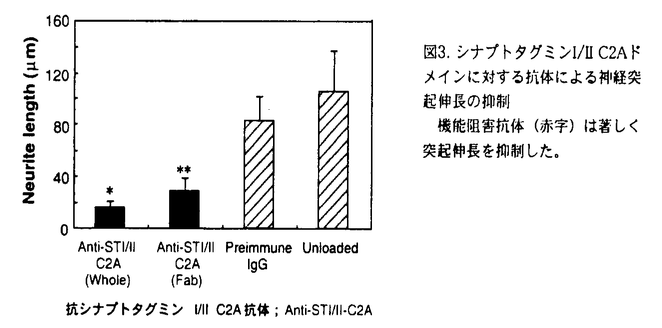 図3. シナプトタグミンI/IIC2Aドメインに対する抗体による神経突起伸長の抑制機能阻害抗体(赤字)は著しく突起伸長を抑制した。 図3. シナプトタグミンI/IIC2Aドメインに対する抗体による神経突起伸長の抑制機能阻害抗体(赤字)は著しく突起伸長を抑制した。 2,マウス・シナプトタグミンI/IIのC2Aドメインに対する特異的な抗体遺伝子(EGFP-A36)をエレクトロポーレーション法により、マウス16日胚のDRG神経細胞に導入し、突起伸長への影響を調べた。その結果、発現した抗体は神経突起の数には影響を与えなかったが、突起伸長を著しく阻害した(図4)。 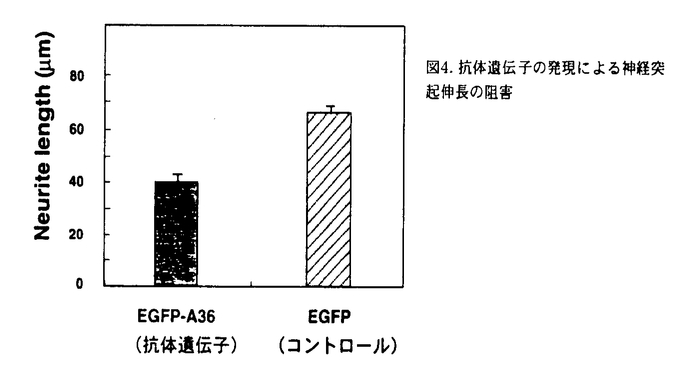 図4. 抗体遺伝子の発現による神経突起伸長の阻害 図4. 抗体遺伝子の発現による神経突起伸長の阻害 以上の結果から、シナプトタグミンI/IIのC2Aドメインは神経突起伸長にも関与する事が明らかとなった。また、形成される神経突起の数には影響がなかったことから、シナプトタグミンI/IIのC2Aドメインは神経突起の発芽には関与しない可能性が示唆された。 本研究で使用した機能阻害抗体(anti-STI/II-C2A)はC2AドメインのCa2+依存的なリン脂質やシンタキシンとの結合をin vitroで阻害し、シナプス小胞とシナプス前膜との融合過程をin vivoで阻害することが分かっている。このことからシナプス小胞とシナプス前膜との融合過程にはシナプトタグミンのC2AドメインがCa2+依存的にリン脂質やシンタキシンと結合することが重要と考えられている。従って、本実験で得られた神経突起伸長における抗体の阻害効果も、成長円錐小胞と成長円錐形質膜の融合過程を抑制した結果である可能性が高い。また、シンタキシンは神経突起伸長に関与することが既に報告されており、その突起伸長時における分布もシナプトタグミンと非常に良く似ている。従ってシナプトタグミンとシンタキシンのCa2+依存的な結合が神経突起伸長にも重要な役割をはたしているものと推測される。 |