本研究は、放射線等によるDNA損傷に対する応答や染色体不安定性,種々の成人性疾病との関連が示唆されているpoly(ADP-ribosyl)化の細胞内での役割を明らかにするために、poly(ADP-ribose)の分解酵素であるParg(poly(ADP-ribose)glycohydrolase)の精製・遺伝子クローニングおよび遺伝子座の決定を試みたものであり、下記の結果を得ている。 1.Poly(ADP-ribose)をリガンドとして用いたアフィニティカラムを含む6種類のカラムにより、ラット精巣よりpoly(ADP-ribose)分解活性を指標としてPargタンパク質を精製した。約1万倍に精製された最終精製画分中に含まれる複数のタンパク質より、改良を加えた1次元zymogram法を用いて60kDaのタンパク質がPargであることを示した。 2.精製した60kDa Pargの部分アミノ酸配列を決定した。その配列に基づいてdegenerated primerを作製し、RT-PCR法と5’RACE法により全長3,974bpのラットParg cDNAを単離し、全塩基配列を決定した。単離したラットParg cDNAは972アミノ酸からなる推定分子量109kDaのタンパク質をコードしていることを示した。 3.単離したcDNAがコードするタンパク質をGST融合タンパク質として発現させ、この109kDaのタンパク質にParg活性があること、C末端部分にpoly(ADP-ribose)分解を担う領域があることを示した。 4.単離したParg cDNA断片をプローブとしてノーザンブロット解析を行い、ラットParg mRNAが解析したすべての組織で約4kbの1種類だけが発現していることを示した。 5.ラットPargイントロン配列中に、F344ラットとACIラットの間で遺伝多型が存在することを見出した。さらにこの遺伝多型についてPCR-SSCP法により139匹の(F344xACI)F1xACIバッククロスラットのタイピングを行い、遺伝多型マーカーとの連鎖解析により、ラットParg遺伝子を第16番染色体短腕上、D16Mit2マーカーと同じ位置にマップした。 6.ヒトPARGゲノムDNAを含むP1クローンを単離した。このクローンをプローブとして、FISH法によりヒトPARG遺伝子を第10番染色体q11.23-21.1にマップした。 以上、本論文はpoly(ADP-ribose)の分解酵素について、タンパク質の精製を経て、重要な基礎的知見(塩基配列、遺伝子座)を明らかにした。本研究により、これまでほとんど解析出来なかったpoly(ADP-ribose)の分解制御機構について分子生物学的手法を用いた研究ができるようになり、DNA損傷応答などにおけるpoly(ADP-ribosyl)化の役割の解明に重要な貢献をなすと考えられ、学位の授与に値するものと考えられる。 | 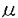 gを得た。最終精製画分をSDS-ポリアクリルアミドゲルで電気泳動した後、銀染色したところ、複数のタンパク質が検出された。ゲル中で活性測定を行うl次元zymogram法により、60kDaのタンパク質にのみpoly(ADP-ribose)分解活性が確認されたことから、この60kDaのタンパク質が目的のPargであると結論した。部分精製したPargは、pl値は8.0〜8.7、平均鎖長22ADP-riboseのpoly(ADP-ribose)に対するKm値は3.2
gを得た。最終精製画分をSDS-ポリアクリルアミドゲルで電気泳動した後、銀染色したところ、複数のタンパク質が検出された。ゲル中で活性測定を行うl次元zymogram法により、60kDaのタンパク質にのみpoly(ADP-ribose)分解活性が確認されたことから、この60kDaのタンパク質が目的のPargであると結論した。部分精製したPargは、pl値は8.0〜8.7、平均鎖長22ADP-riboseのpoly(ADP-ribose)に対するKm値は3.2