低密度リポタンパク質受容体(LDLR)はアポリポタンパク質BまたはE(アポBまたはアポE)を含むリポタンパク質(LDL,IDL,VLDL等)をリガンドとして結合し、リガンドと共に細胞内に取り込まれる細胞膜上の受容体である。その機能異常はヒトでは家族性高コレステロール血症の原因となる。 LDLRのファミリーにはそのほか、哺乳類やニワトリからクローニングされた超低密度リポタンパク質受容体(VLDLR)、アポタンパク質E受容体2(apoER2)、リポタンパク質受容体関連タンパク質1(LRP1)、LRP2、LRP3、LRP5(LRP7,LR3)、LRP6、ST7およびLR11/SorLAが現在までに知られている。ショウジョウバエのY1(yorkless)や線虫Caenorhaditis elegansのLRPもこのファミリーに含まれる。これらの蛋白質の共通の特徴はLDLRのリガンド結合ドメイン(1igand binding repeat,LBR)に相同のドメインを複数持つI型膜タンパク質であることである。 この論文では、マウスリンパ球からLDLRファミリーの新規タンパク質をcDNAクローニングしたので報告する。 筆者は、既に報告されているレトロウイルスベクターを用いたシグナルシークエンストラップ法を用い、マウスリンパ球のcDNAライブラリからI型膜タンパク質または分泌タンパク質をスクリーニングすることによって、未知のクローンを得た。この全長をコードするcDNAクローンを再度スクリーニングし塩基配列を決定したところ、2142塩基のオープンリーディングフレームを持っていた。17アミノ酸の典型的シグナルシークエンスがあり、成熟した産物は696アミノ酸からなる計算上の分子量が74.8kDaのI型膜蛋白であることがわかった。遺伝子バンクのホモロジー検索の結果、LDLRのLBRに相同の部位を持つ新規のLDLR関連タンパク質であった。これをLDL receptor-related protein 9(LRP9)と名付けた。 細胞外部分は4つのLBRと2つのCUBドメインと呼ばれる構造からなる。 LDLRのLBRにある酸性残基はリガンドとの結合に必要であるが、LRP9においてもこの酸性残基は高度に保存されている。LRP9もLBRを介して何らかのリガンドが結合すると考えられる。 CUBドメインは補体C1s/C1r、BMP-1などに共通に見られる弱く保存された配列を呼ぶ。その機能的特徴は未だ不明である。従ってLRP9のCUBドメインの機能も不明である。 細胞外ドメインはアミノ末端から順に一つのCUBドメイン、一つのLBR、一つのCUBドメイン、三つのLBRと並ぶ。LDLR・VLDLRが持つO-糖鎖結合配列や上皮性成長因子(EGF)様ドメインはLRP9にはない。EGF様ドメインは、LDLRにおいては酸性依存性のリガンドとの解離に働くと考えられている。 一回の膜貫通部分を経て、細胞内にはPPSYモチーフとプロリンリッチ領域がある。LDLRにおいてNPXYモチーフがリガンド・受容体複合体の細胞内への取り込みに働くとされている。また、VLDLRとapoER2は神経遊走に必要なmDab1シグナルの直接の上流に位置すると考えられているが、mDab1はVLDLR-apoER2のNPXYモチーフに結合するというin vitroの知見がある。LRP9においてはNPXYモチーフはなく、PPSYモチーフがある。これはLRP9、LRP3、ST7に共通した配列で何らかの生理的役割を果たしている可能性がある。 ラディエーションハイブリッドによりLRP9をコードする遺伝子はマウス染色体14番上にマップされた。マウスLRP9に極めて相同性の高いヒトESTクローンが報告されており、ヒトLRP9 cDNAの部分配列と考えられることから、この配列をもとにヒトLRP9をコードする遺伝子座を決定したところ、14q11.2にマップされた。 次に、ノーザンブロットにより発現分布をmRNAレベルで解析した。マウスにおいては心臓、肺、腎臓、肝臓において比較的強く、また脳と脾臓において比較的弱く発現が認められた。骨格筋と精巣における発現は検出限界以下であった。マウス胎児においては、胎生7日で強い発現を示した後、胎生11日で発現が弱まり、その後胎生15日・17日と発現が再び増加した。 マウスのcDNAプローブを用いたヒトでのノーザンブロットの結果はマウスとは異なっており、発現分布は広く末梢白血球、肺、胎盤、小腸、肝臓、腎臓、脾臓、胸腺、大腸、骨格筋、心臓において発現が認められた。脳においては発現を認めなかった。 アダルトマウスの脳でin situハイブリダイゼーションを行ったところ、choroid plexus・ 第三脳室上衣細胞・海馬CA3領域・軟膜に限局して発現が見られた。 LRP9はLDLRのLBRと相同性の領域を持つので、既知のリポ蛋白をラベルし、これをLRP9が認識するかどうかを調べた。 まず初めに、CHO細胞の亜株で内因性のLDLRを欠損したldl-A7株に、LRP9 cDNAを発現させた。これをクローン化し、LRP9の発現量の多いクローンを以下の実験に用いた(ldl-A7/LRP9)。 細胞を蛍光(DiI:1,1’-dioctadecyl-3,3,3’3’-tetramethyl indocarbocyanine perchlorate)ラベルしたしLDLとインキュベートすると、野生型のCHOはLDLを取り込んだがldl-A7/LRP9、ldl-A7/mockとldl-A7はこれを取り込まなかった。アセチルLDL(AC-LDL)も取り込まなかった。従って、LDLとAc-LDLはLRP9のリガンドではないと考えられた。 次に、放射能(125I)ラベルしたVLDL、LDL、HDL及びAc-LDLを培養細胞が取り込み、代謝して培養上清中に放出する125I-チロシンの放射活性を測定するデグラデーションアッセイを行った。ldl-A7/mockやldl-A7に比し、ldl-A7/LRP9はいずれのリガンドも優位にデグラデートしなかった。VLDL,LDL,HDL及びAc-LDLはLRP9のリガンドではないと考えられた。 カイロミクロンレムナントが肝細胞に取り込まれる際、肝細胞表面にあるアポEが付加されてアポEリッチなりポタンパク質が局所的に形成され、受容体への親和性を増して取り込まれる経路があると考えられている。このリポタンパク質は実験上はウサギにコレステロール負荷をした際に現れる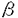 -VLDLで代用される。 -VLDLで代用される。 -VLDLのバインディングアッセイの結果、ldl-A7/LRP9は20-50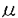 g/mlのリガンド濃度において親株に比し有意に-VLDLを結合した。従ってLRP9はアポEの低親和性受容体である可能性がある。しかしこの実験においては50g/mlのリガンド濃度においてもリガンド結合は飽和していない。結合の特異性を確認するためにさらに高いリガンド濃度において結合が飽和することを示す必要がある。更に、-VLDLの結合や取り込みを確認するためリガンドブロットやコレステリル・エステリフィケーション・アッセイを行っている。-VLDLとLRP9の結合の生理的意義に関しても今後の検討が必要である。 g/mlのリガンド濃度において親株に比し有意に-VLDLを結合した。従ってLRP9はアポEの低親和性受容体である可能性がある。しかしこの実験においては50g/mlのリガンド濃度においてもリガンド結合は飽和していない。結合の特異性を確認するためにさらに高いリガンド濃度において結合が飽和することを示す必要がある。更に、-VLDLの結合や取り込みを確認するためリガンドブロットやコレステリル・エステリフィケーション・アッセイを行っている。-VLDLとLRP9の結合の生理的意義に関しても今後の検討が必要である。 LRP9のドメインの構造はLDLRとは異なる特徴を持つ。LRP3・ST7もこの特徴を共有し、これらはLDLRファミリー中のサブファミリーを形成すると考えられる。アミノ酸配列上のホモロジーはLRP9とLRP3・LRP9とST7のともに約50%である。 しかしながら、その発現分布はLRP3・ST7とは全く異なる。肝臓・腎臓においてLRP9は強く発現しているが、LRP3・ST7は発現を認めない。従ってLRP9の生理的役割もLRP3・ST7とは異なると考えられる。構造的に類似する二つのタンパク質が存在すること、マウスLRP9クローンとヒトESTクローンの配列が高度に保存されていること、LRP9がブラストシスト期から発現していること(LRP9 cDNAの部分配列に一致するブラストシスト由来のESTクローンが報告されている)は、LRP9が生理的に重要な役割を果たしていることの傍証となる。 LRP9が発現している臓器は血管に富む臓器か、または上皮由来の臓器と考えることができる。脳におけるin situハイブリダイゼーションの結果はこの考えに矛盾しない。 レムナントの肝臓での取り込みについては不明な点が残されている。即ちLDLRが欠損する家族性高コレステロール血症の肝臓でもレムナントを正常に取り込むという事実である。従って肝臓における第二のレムナント受容体があると予想されている。-VLDLのバインディングアッセイの結果は、LRP9がマウスの肝臓におけるカイロミクロンレムナントに対する新しい低親和性受容体である可能性を示す。 LRP9の脳での発現部位はLRP2に似る。LRP2を欠損するマウスはholoprosencephalic syndromeを示す。また腎臓においてはビタミンD代謝物の再吸収ができなくなり骨形成不全を来す。LRP9は腎臓でも発現しており、LRP9のこれらの臓器における生理的役割を発生工学的に明らかにすることは興味深い。 ヒトlrp9遺伝子がmapされた14q11.2の近傍に関連する原因遺伝子不明の遺伝性疾患としてdistal myopathiesとspastic paraplegiaがある。これら疾患とLRP9との関連はここでは調べなかった。 この論文においては、新規LDLR関連タンパク質であるマウスLRP9のcDNAクローニングを行った。LRP9にはLDLRファミリーに共通するLBRが保存され、細胞外領域に4回繰り返していた。マウスにおいては心臓、肺、腎臓、肝臓において比較的強く、また脳と脾臓において比較的弱く発現が認められた。遺伝子は染色体14番に位置する。LDL・VLDL・HDL・Ac-LDLはLRP9のリガンドではなかった。LRP9を強制発現したldl-A7細胞はin vitroで-VLDLと低親和性に結合した。LRP9の生理的機能を解明するために、-VLDLとの結合のさらなる解析、腎臓・肝臓における解剖学的局在の解明を現在行っている。 |