| 研究の背景および目的 APC(adenomatous polyposis coli)遺伝子は、家族性大腸腺腫症の家系の胚細胞や大部分の散発性大腸ポリープや大腸癌で変異を認めることが知られている。また、APCタンパク質は、細胞接着にも重要な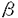 -cateninと結合し、その分解を誘導することによって、転写因子Tcf(T ce11 factor)を介したシグナルを負に制御している。正常な上皮細胞においては、APC/-catenin複合体はGSK-3(glycogen synthase kinase)によりリン酸化され、リン酸化された-cateninは速やかに分解される。変異型APCは-cateninとの複合体を形成できず、その分解を妨げ、その結果細胞質内で安定化した-cateninはTcfと結合し、核内へ移行し、c-mycやcyclin D1といった標的遺伝子の転写活性化を引き起こし、癌化に寄与すると考えられている。 -cateninと結合し、その分解を誘導することによって、転写因子Tcf(T ce11 factor)を介したシグナルを負に制御している。正常な上皮細胞においては、APC/-catenin複合体はGSK-3(glycogen synthase kinase)によりリン酸化され、リン酸化された-cateninは速やかに分解される。変異型APCは-cateninとの複合体を形成できず、その分解を妨げ、その結果細胞質内で安定化した-cateninはTcfと結合し、核内へ移行し、c-mycやcyclin D1といった標的遺伝子の転写活性化を引き起こし、癌化に寄与すると考えられている。 近年、APCに変異のない大腸癌で、-catenin遺伝子のエクソン3に存在するGSK-3によるリン酸化部位の変異により、-catenin/Tcfシグナルが活性化されることが示され、その後、他の臓器由来の癌でも報告されている。 過去の-catenin/Tcfシグナルの異常についての検討は、APCあるいは-catenin遺伝子の変異に関するものがほとんどであり、様々な臓器由来の癌においてAPCあるいは-catenin遺伝子の変異以外の機序も含めてどの程度の割合で、-catenin/Tcfシグナルの活性化が起こっているかは明らかではない。本研究では、細胞株を用いて、各種消化器癌における-catenin/Tcfシグナルの活性化の頻度とAPCあるいは-cateninの変異の寄与を検討した。 研究方法 ヒト大腸癌細胞株12株、ヒト胃癌細胞株8株、ヒト膵癌細胞株9株、ヒト肝細胞癌細胞株7株の合計36細胞株を用いて実験を行なった。 まず、-catenin/Tcfシグナルのレポータープラスミドとして、Tcf結合配列にルシフェラーゼ遺伝子が連結された構造を有するpTOPFLASH及びTcf結合配列に変異が導入されているpFOPFLASHを各細胞株にトランスフェクションし、24時間後にルシフェラーゼアッセイを行なった。pTOPFLASHとpFOPFLASHのルシフェラーゼ活性の比を、各細胞株の-catenin/Tcfシグナル活性とした。また、-catenin/Tcfシグナルの活性化を-catenin核染色の有無により確認するため、-cateninの免疫染色を行なった。次に、シグナルの活性化を認めた細胞株におけるAPC及び-cateninの変異の有無を、APCの変異はイムノブロット及びエクソン15のmutation cluster regionを標的としたprotein truncation testによるtruncateしたAPC遺伝子産物の検出により、-cateninの変異はエクソン3のGSK-3のリン酸化部位のシークエンスにより、検討した。 結果 大腸癌細胞株では12細胞株すべてでシグナルの活性化を認めたが、それに対し、膵癌細胞株では9細胞株すべてシグナルの活性化は認めず、胃癌細胞株では8細胞株中5細胞株に、肝細胞癌細胞株では7細胞株中2細胞株にシグナルの活性化を認めた。 -catenin/Tcfシグナルの活性化を認めた大腸癌細胞株12株全て、胃癌細胞株5株、肝細胞癌細胞株2株及びシグナルの活性化を認めなかった細胞株のうち胃癌細胞株1株、肝細胞癌細胞株3株、膵癌細胞株3株について、-cateninの免疫染色を行ったところ、-catenin/Tcfシグナルの活性化を認めた19細胞株全てにおいて、-cateninの核内への蓄積が認められたが、活性化を認めなかった7細胞株では-cateninは細胞膜に局在し、核への蓄積は認められなかった。 次に-catenin/Tcfシグナルの活性化が認められた19細胞株(大腸癌12株、胃癌5株、肝細胞癌2株)において、APCあるいは-cateninの変異の関与について検討した。イムノブロット及びprotein truncation testでtruncateしたAPC遺伝子産物を認めたものは大腸癌細胞株CaR-1、CaCO2、Colo320、DLD1、HCT15、LOVO、SW620、SW837、胃癌細胞株MKN28、MKN74の10細胞株であった。一方、シークエンス解析により-catenin遺伝子エクソン3に変異を認めたものは大腸癌細胞株HCT116(コドン45のSerの欠失)、SW48(コドン33のSer(TCT)からTyr(TAT)への置換)、胃癌細胞株AGS(コドン34のGly(GGA)からGlu(GAA)への置換)、肝細胞癌細胞株HuH6(コドン34のGly(GGA)からVal(GTA)への置換)の4細胞株であった。また、肝細胞癌細胞株HepG2はエクソン3欠失が過去の報告にみられたが、今回のシークエンス解析では検出できなかった。大腸癌細胞株RCM1は-cateninには変異を認めなかったが、APCの変異の有無は今回の検討では確認できなかった。残りの3細胞株、大腸癌細胞株LS174T、胃癌細胞株AZ-H6C、MKN45は、APC、-catenin共に変異を認めなかった。 考察 本研究では、さまざまな臓器由来の癌に-catenin/Tcfシグナルの活性化がどのように関わっているかを明らかにするため、胃癌8、大腸癌12、膵癌9及び肝細胞癌7細胞株における-catenin/Tcfシグナルの転写活性を検討した。 大腸癌細胞株では、12株全てにおいて-catenin/Tcfシグナルの活性化を認めた。このような活性化はAPC遺伝子あるいは-catenin遺伝子の変異のある細胞に認められるとの報告がある。そのため、これらの細胞株におけるAPCと-cateninの変異について検討した。12細胞株中APCあるいは-catenin遺伝子の変異を認めたのは10細胞株であったが、APC及び-catenin遺伝子がともに正常である大腸癌細胞株も存在した(LS174T)。このことは大腸癌においてはAPCあるいは-catenin遺伝子の変異の有無に関わらず-catenin/Tcfシグナルが活性化しており、癌化に重要な役割を持っていることを示唆している。 大腸癌とは対照的に、膵癌9細胞株においては-catenin/Tcfシグナルの活性化は全く認められず、-catenin/Tcfシグナルの活性化は膵癌にはほとんど関係ないと考えられる。 胃癌細胞株においては8細胞株中5細胞株(63%)に-catenin/Tcfシグナルの活性化が認められ、これら5細胞株(AGS、AZ-H6c、MKN28、MKN45及びMKN74)おけるAPC遺伝子及び-catenin遺伝子エクソン3の変異を調べたところ、MKN28及びMKN74細胞にAPCの変異を、AGS細胞に-catenin遺伝子の変異を認めた。しかし、AZ-H6C及びMKN45にはAPCにも-cateninエクソン3にも変異を認めなかった。この結果、これら2つの細胞株では大腸癌細胞株LS174Tと同様に、APCあるいは-catenin変異以外の-catenin/Tcfシグナル活性化の機序が存在することが示唆される。本研究の結果は-catenin/Tcfシグナル活性化はかなり多くの胃癌において重要な働きを有し、この活性化はAPCあるいは-cateninの変異によってのみ引き起こされるものではないことを示唆している。消化管由来の癌である大腸癌と胃癌には共通の癌化のメカニズムがあることが推察される。 肝細胞癌では約20%に-catenin遺伝子の変異を認めるという報告があるが、APC遺伝子変異の報告はほとんどない。本研究では肝細胞癌7細胞株における-catenin/Tcfシグナル活性を調べたところ、HepG2及びHuH6の2細胞株にのみシグナルの活性化を認めた。これらは共に-catenin遺伝子エクソン3の変異を認める細胞株であったことから、-catenin/Tcfシグナルはある種の肝細胞癌でのみ重要な働きを持ち、そのシグナル活性化機序は主として-cateninの変異によることが示唆された。 結論 -catenin/Tcfシグナルの活性化は、大腸癌12細胞株全て(100%)に認められ、胃癌でも8細胞株中5細胞株(63%)に認められたが、肝細胞癌では7細胞株中2細胞株(29%)に認められたのみであり、膵癌9細胞株には全く認められなかった(0%)。このことは、消化管粘膜由来の癌において、-catenin/Tcfシグナルの活性化が重要な働きを担っているのに対し、肝臓由来の癌においては重要性は低く、膵臓由来の癌においてはほとんど重要な働きを持たないことを示唆している。 また、-catenin/Tcfシグナルの活性化機序は主として、APC遺伝子あるいは-catenin遺伝子の変異であったが、APC遺伝子にも-catenin遺伝子にも変異を持たない胃癌や大腸癌細胞株の中に、-catenin/Tcfシグナルが活性化された細胞株が認められたことから、-catenin/Tcfシグナル活性化の他の機序が存在することが示唆された。 |  -catenin/T cell factorシグナル伝達異常の解析
-catenin/T cell factorシグナル伝達異常の解析