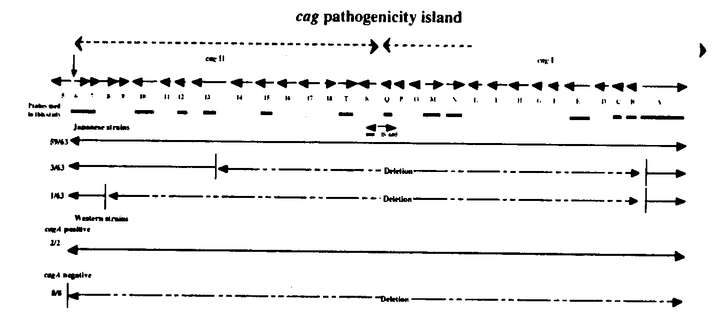
本研究はHelicobacter pyloriの病原因子とされるPathogenicity island(PAI)の構造およびその機能を明らかにする目的で、PAIに存在するCagAに対する抗体、蛋白の検出、およびPAI遺伝子群の検出系の確立を行い、多数例において検討した。加えて、PAIの機能を解明する目的で、NF-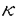 B活性化機構について検討を行ったものであり、下記の結果を得ている。 B活性化機構について検討を行ったものであり、下記の結果を得ている。 1. H.pylori標準株であるATCC43526のcagA遺伝子全長をクローニングし、全塩基配列を決定した。アミノ酸配列で比較すると、既報の2株とは90%以上の相同性を認めたが、後半部分には繰り返し構造が3回存在し、200から300塩基長い遺伝子であった。 2. H.pylori cagA遺伝子の一部(1344bp)を蛋白発現用プラスミドベクターに組換え、E.coli JM109株に導入し、CagA蛋白の発現を誘導・精製した。H.pylori株中のCagA蛋白の存在を組換え蛋白に対して作製した家兎抗血清をもちいてイムノブロット法にて検討した。また、内視鏡検査を施行した患者血清中の抗CagA抗体の存在を組換えCagA蛋白を抗原としたイムノブロット法を用いて調べた。その結果血清抗体、菌内蛋白ともにわが国では90%以上の陽性率であり、患者の疾患間に差を認めなかった。 3. 約30のPAI遺伝子のうち13の遺伝子をクローニングし、それらをプローブとして63株の臨床分離株におけるPAIの構造を検討した。その結果59株は完全なPAIを有し、4株に部分欠損が認められた。これまでcag PAIのマーカーと考えられていたcagA遺伝子は全株に存在した。これらの株についてCagA蛋白、およびTranscriptとの関係を調べたところ、これらの株はcagA遺伝子が存在するにもかかわらず、Transcriptおよび蛋白が欠損していた。加えて、これらの株は胃癌細胞株からのIL-8誘導能が欠損していた。 4. H.pyloriによるIL-8誘導には転写因子としてNF-Bが関与している。その経路解析のためH.pyloriのNF-B活性化をNF-B-Luc(5xNF-B site)を用いたルシフェラーゼアッセイにて検討した。その結果cag PAI陽性のTN2、26695株はNF-Bを活性化したが、部分欠損株、完全欠損株およびTN2- cagEでは活性化は見られなかった。IB cagEでは活性化は見られなかった。IB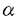 の活性化を、H.pyloriを共培養した細胞(MKN45)より蛋白を抽出し、抗IB抗体を用いたイムノブロットによる、IB蛋白量の変化、およびIBのDominant negativeを発現させ、NF-Bの活性化の変化により検討したところ、イムノブロットではH.pyloriにてdegradationが観察され、さらにDominant negativeによってNF-Bの活性化は抑制された。IBのキナーゼであるIKKおよびIKK の活性化を、H.pyloriを共培養した細胞(MKN45)より蛋白を抽出し、抗IB抗体を用いたイムノブロットによる、IB蛋白量の変化、およびIBのDominant negativeを発現させ、NF-Bの活性化の変化により検討したところ、イムノブロットではH.pyloriにてdegradationが観察され、さらにDominant negativeによってNF-Bの活性化は抑制された。IBのキナーゼであるIKKおよびIKK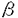 とH.pyloriとの関連をFLAGラベルされたIKK、IKKを蛋白発現させ、抗FLAG抗体で免疫沈降後、GST-IBを基質としてキナーゼアッセイを行った。また、IKKのDominant negative(IKK(K44M)およびIKK(K44M))を発現させ、H.pyloriによる変化を観察した。その結果、H.pyloriとの共培養によりIKKの活性化が観察された。また、IKKのDominant negativeによりNF-Bの活性化は容量依存性に抑制された。これらのことより、H.pyloriはIKK、IKKを活性化させてNF-Bを活性化することが示された。IKKの上流シグナルの検討としてNIK、TRAF2、TRAF6のDominant negativeによる影響を検討したところ、NIKおよびTRAF2とTRAF6のDominant negativeによって、NF-Bの活性化は容量依存性の抑制を示し、NIK、TRAF2およびTRAF6がH.pyloriのNF-B活性化に関与していることが示された。 とH.pyloriとの関連をFLAGラベルされたIKK、IKKを蛋白発現させ、抗FLAG抗体で免疫沈降後、GST-IBを基質としてキナーゼアッセイを行った。また、IKKのDominant negative(IKK(K44M)およびIKK(K44M))を発現させ、H.pyloriによる変化を観察した。その結果、H.pyloriとの共培養によりIKKの活性化が観察された。また、IKKのDominant negativeによりNF-Bの活性化は容量依存性に抑制された。これらのことより、H.pyloriはIKK、IKKを活性化させてNF-Bを活性化することが示された。IKKの上流シグナルの検討としてNIK、TRAF2、TRAF6のDominant negativeによる影響を検討したところ、NIKおよびTRAF2とTRAF6のDominant negativeによって、NF-Bの活性化は容量依存性の抑制を示し、NIK、TRAF2およびTRAF6がH.pyloriのNF-B活性化に関与していることが示された。 以上、本論文はH.pyloriの病原因子であるcag pathogenicity islandについて、その構造解析の結果、日本の臨床分離株のほとんどがintactなcag PAI陽性株であることを示し、またその病原性を既定していると考えられるNF-B活性化経路について膜近傍の因子までの関与を示唆した初めての論文であり、学位の授与に値すると考えられる。 |