本研究は骨形成において中心的な役割を果たしている骨芽細胞の増殖と分化における刀子レベルでの制御機構を検討するために、その最終決定機構である細胞周期の制御因子、特に細胞周期進行の負の制御因子であるCDKインヒビターの蛋白質レベルでの関与を解明することを試みたものであり下記の結果を得ている。 I.ラット初代培養骨芽細胞においてCip/Kipファミリーであるp27、p57、p21の3種類の蛋白はすべて発現していた。そしてこの細胞に、TGF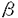 1とBMP-7を添加した時、増殖は共に促進された。ところが、分化に関連して、TGF1投与群では非添加群に比してALP活性は抑制され、逆にBMP投与群ではALP活性が上昇した。これら細胞においてCip/Kipファミリーの発現は、TGF1投与群ではp57の発現が劇的に減少した。これに対して、BMP-7投与群では発現に著明な変化はなかった。 1とBMP-7を添加した時、増殖は共に促進された。ところが、分化に関連して、TGF1投与群では非添加群に比してALP活性は抑制され、逆にBMP投与群ではALP活性が上昇した。これら細胞においてCip/Kipファミリーの発現は、TGF1投与群ではp57の発現が劇的に減少した。これに対して、BMP-7投与群では発現に著明な変化はなかった。 II.CDK2とp57の結合は非添加群ではその結合が確認出来るが、TGF1刺激時では消失していること、CDK2の酵素活性が上昇していること、ならびにp107の発現と上方へのバンドシフトが確認された。 III.TGF1によるp57の劇的な減少はプロテアソーム阻害剤により著明に抑制されること、そしてp57蛋白質と単数さらには複数のGST-ユビキチンがin vitroで結合することを明らかにし、TGF1添加によるp57の発現減少がユビキチン-プロテアソームによる蛋白分解機構の活性化によることを証明した。 IV.BMP-7添加時には、CDK2とp27もしくはp57の結合は非添加時と比て変化せず、結合したままであった。しかし、CDK2の酵素活性は上昇し、p107の上方へのバンドシフトが確認された。さらにBMP-7添加時のサイクリンの発現を確認したところ、非添加時と比較してサイクリンD1、D2、E、Aの発現が誘導され、さらにこの発現誘導はTGF1刺激と比較しても強力であることが示された。 以上、本研究では、骨芽細胞の増殖と分化の制御における細胞周期制御因子の関与を明らかにした。さらに骨基質に大量に存在するサイトカインであるTGFによる骨芽細胞の増殖と分化の制御においてはCDKインヒビターの一つである、p57が重要な役割を果たしていることが示された。 本研究は骨芽細胞の増殖と分化、即ち、骨形成における分子レベルでの制御機構の解明に役立つと考えられ、学位の授与に値するものと考えられる。 |