| 要約 CD23はIgEの低親和性の受容体であり、さまざまな骨髄由来の細胞においてその発現が確認されている。ヒト表皮ランゲルハンス細胞(以下LC)においてもCD23の発現は確認されているが、マウスLCでは発現していないとされている。我々は抗Ia抗体を用いたパンニング法により、マウスの表皮から95%以上の純度でLCを採取することが可能になったので、この方法によってマウスLCでのCD23発現について検討することとした。その結果、蛋白レベル、mRNAのいずれによっても培養LCにおいてその発現を認めた。LCと脾臓より得た白血球のCD23の塩基配列を比較したところほぼ同一の配列が認められた。CD23のLCにおける発現はケラチノサイト(以下KC)由来のサイトカインであるIL-1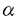 、IL-18、GM-CSF、M-CSF、TNF-によって抑制を受けた。また,抗CD23抗体(クローンB3B4)により、培養LCによるIL-6産生が抑制されたが、IL-10、TNF-については抑制はみられなかった。さらにLCはIgEを結合することが明らかとなり、この結合は部分的に抗CD23抗体によって抑制されることも明らかになった。以上によってマウスLCはCD23を発現し、そのCD23を介して免疫反応に関与することが示唆された。また、これまでマウスLCでCD23の発現がないとされてきた主な理由はKC由来のサイトカインによってその発現が抑制されていたためと考えた。 、IL-18、GM-CSF、M-CSF、TNF-によって抑制を受けた。また,抗CD23抗体(クローンB3B4)により、培養LCによるIL-6産生が抑制されたが、IL-10、TNF-については抑制はみられなかった。さらにLCはIgEを結合することが明らかとなり、この結合は部分的に抗CD23抗体によって抑制されることも明らかになった。以上によってマウスLCはCD23を発現し、そのCD23を介して免疫反応に関与することが示唆された。また、これまでマウスLCでCD23の発現がないとされてきた主な理由はKC由来のサイトカインによってその発現が抑制されていたためと考えた。 研究目的 IgEの低親和性のレセプターであるCD23はアトピー性皮膚炎患者で発現が亢進していることが知られており、慢性炎症性疾患に関係していると考えられている。CD23はB細胞、T細胞、好酸球、マクロファージ、肥満細胞、血小板、単球、ランゲルハンス細胞など造血系の細胞以外に、KCにも発現が確認されている。 LCは表皮に存在する骨髄由来のMHCクラスII抗原陽性の抗原提示細胞であり、全表皮の1-3%を占める。ヒトめ表皮ではCD23はLC、KCいずれにおいてもその発現が確認されているが、マウスの表皮ではKCもLCもCD23を発現しないとされている。 これまでマウスLCに関する研究はKCが多く混合した比較的純度の低い実験系に関するものであるので、マウスLCにおいてCD23の発現が確認できなかった理由としてKC由来のサイトカインの影響を考え、95%以上の純度で純化したマウスLCを用いて、CD23の発現について再評価を行った。また、LC上のCD23の機能についても検討を加えた。 方法 実験1 Balb/cマウスの躯幹の皮膚を採取し、ディスパーゼ処理をし、表皮を剥離した後、DNase処理を加え、表皮細胞浮遊液を作製した。その後、抗Ia抗体と反応させ、予めヤギ抗マウスIgG抗体を付着させておいたパンニングディッシュ上に付着させ、付着細胞を回収し精製新鮮LC(fLC)とした。非付着細胞はLC除去表皮細胞(LC-depleted EC)とした。これらの細胞のCD23の発現を培養前後で比較した。 実験2 上記により採取したLCを培養し、培養LC(cLC)よりmRNAを採取し、cDNAを合成し、CD23に対するプライマーを用いてRT-PCRおよびnested PCR法によりcLCおよびLC-depleted ECのCD23の発現を検討した。また、nested PCR産物のDNA塩基配列を決定し、すでに決定されているCD23の全エクソンと比較した。 実験3 fLCにKC由来のサイトカイン(IL-1、IL-10、IL-18、TNF-、M-CSF、GM-CSF)を加えて培養しcLC上のCD23の発現の変化をFACSを用いて検討した。 実験4 fLCを24時間培養後、抗マウスCD23抗体(B3B4)を加えさらに24時間培養した後に、培養上清を回収しELISA法によりIL-6の濃度を測定した。また、cLC上に発現したCD23にIgEが結合できるかも検討した。 実験5 アトピー性皮膚炎のモデルマウスとされるNC/Ngaマウスの皮膚の凍結切片を用いて、免疫染色を行い、表皮のランゲルハンス細胞のCD23の発現について検討した。 結果実験1蛋白レベルでのCD23の発現 fLCあるいはcLCをFITC標識抗マウスCD23抗体で染色し、CD23のMFIを測定し分析した。その結果、cLCのCD23の発現は新鮮LCと比較して時間依存性に有意に増加していた。また、FITC標識およびPE標識モノクロナール抗体を用いた2カラー染色によりCD23の発現はIa抗原陽性細胞に限られており、LC-depleted ECではCD23の発現は蛋白レベルでは認められなかった。 実験2mRNAレベルでのCD23の発現 CD23の発現をmRNAレベルで検討した結果、cLCより作製したcDNAよりRT-PCRおよびnested PCR法によりCD23の発現を確認できた。nested PCR産物のDNAの塩基配列を決定し、対応するマウスのCD23のエクソンと比較した結果、数カ所のヌクレオチドの違いがみられただけであった。また、LC-depleted ECより作製したcDNAからはCD23に対応するメッセージは確認できなかった。 実験3KC由来のサイトカインによるCD23発現に対する影響 IL-1、IL-10、IL-18、TNF-、M-CSF、GM-CSFは0.1ng/mlの濃度でcLCのCD23の発現を抑制した。 実験4cLC上のCD23の機能 抗マウスCD23抗体(B3B4)はcLCによるIL-6産生を有意に抑制した。また、cLC上のCD23はIgEを結合でき、B3B4によりこのlgEの結合は有意に抑制された。 実験5 NC/Ngaマウスの表皮LCのin vivoでのCD23の発現 皮膚炎を発症したNC/Ngaマウスの表皮にCD23陽性細胞が確認でき、このCD23陽性細胞はIa抗原陽性であったのでNC/Ngaマウスの表皮LCはin vivoでCD23を発現すると結論した。 考按 マウスの表皮KCは様々なサイトカインの供給源であり、IL-1、IL-6、IL-7、IL-10、IL-18、TNF-、M-CSF、GM-CSFなどを産生することが明らかになっている。これらのサイトカインの多くがcLCのCD23の発現を抑制したので、in vivoの表皮内のLCがCD23を容易に発現できないと考えられる。また、cLCはin vivoで所属リンパ節に求心性に移動した活性化LCに形態、機能の面で類似していると考えられている。CD23はfLCには発現せず、cLCに時間依存性に発現したことから、CD23はin vivoで活性化されたLCの機能に密接に関係していると考えられる。 活性化LC上のCD23がマウスの所属リンパ節や皮膚で機能しているとすれば、2つの役割が考えられる。一つはIgEのレセプターとしての機能であり、もう一つはリガンドとしての機能である。ヒトの場合B細胞上のCD23はIgE免疫複合体のレセプターとして機能し,効率よく抗原をT細胞に提示することが確認されている。我々の実験系でもIgEがCD23に結合することが確認されたので,マウスのLCでも同様のことが皮膚やリンパ節内で起こっている可能性がある。 また,cLC細胞は時間とともにclusterを形成するが,このclusterの形成にCD23がリガンドとして機能している可能性がある。マウスにおいてもヒトにおいてもリンパ節内でのこのようなclusterの形成はまだ確認されていないが,CD23を発現しているLCとCD21などのリガンドをもったT細胞やB細胞がリンパ節内で相互作用している可能性も考えられる。 我々の実験系ではB3B4を加えるとLCによるIL-6の産生が有意に減少した。LCはIL-6を産生し、IL-6は様々な炎症反応に関係することが知られている。現在のところB3B4はCD23への結合を通じて,細胞内にシグナルを送るという考え方が有力なので,B3B4がCD23へ結合することによりIL-6の産生に対して負の調節機能が働き、その結果IL-6の産生が減少すると考えられる。LCのCD23はIL-6の産生の調節を通じて炎症反応の調節に関係している可能性がある。 結論であるが、マウスのLCもヒトのLCと同様、CD23を発現しており、皮膚における様々な免疫反応に関係している可能性が示された。我々のデータは、LCのCD23が関係するヒトにおける諸々の炎症反応の免疫学的、生化学的、細胞学的メカニズムをマウスの系を使って間接的に解明するための実験的戦略を新たに供給した。 |  R II/CD23)on murine Langerhans cells.
R II/CD23)on murine Langerhans cells.