本研究は細菌感染や異物侵入の初期段階において重要な役割を担っている白血球の活性化過程におけるリソソーム酵素放出に着目し、第一に上皮組織中のリソソーム酵素防御物質の探索、第二に白血球で産生される炎症やアレルギー反応の主要なメディエーターであるロイコトリエンB4(LTB4)や血小板活性化因子(PAF)による細胞内におけるリソソーム酵素放出の調節機構を研究したものであり、下記の結果を得ている。 1.消化管上皮組織の脂質抽出を行ったところ、消化酵素が活性化され分泌される十二指腸から回腸において硫酸化脂質コレステロール硫酸が高濃度に存在することが分かった。膵エラスターゼにコレステロール硫酸を添加し、様々な基質を用いて酵素反応を測定したところ、コレステロール硫酸の用量依存性に酵素活性は阻害された。脱硫酸した脂質及びステロイド硫酸やガングリオシドの添加によって酵素活性は影響されず、阻害活性の発現には硫酸基と共に疎水性側鎖が必要であることが分かった。消化管上皮においてコレステロール硫酸は膵エラスターゼ酵素活性を阻害することにより、酵素による自己融解から組織を保護している可能性が見出された。 2.好中球エラスターゼに対するコレステロール硫酸の影響を同様に調べたところ、コレステロール硫酸の用量依存性に酵素活性は阻害された。膵エラスターゼにおける結果と同様に、脱硫酸した脂質及びステロイド硫酸やガングリオシドの添加によって酵素活性は影響されなかった。 3.全身麻酔中の気管支肺胞洗浄液中の脂質抽出を行ったところコレステロール硫酸が検出された。同時に好中球エラスターゼ濃度を測定したところ、麻酔時間の経過とともに上昇していた。一方、コレステロール硫酸合成を合成するコレステロール硫酸基転移酵素活性は、麻酔後1時間または2時間後に活性のピークが認められ、気管支肺胞洗浄液中に分泌されていることが示された。 4.濃縮した気管支肺胞洗浄液上清と抗好中球エラスターゼ抗体との免疫沈降液中に、コレステロール硫酸が検出され、エラスターゼに結合していることが示された。 気管支上皮組織においてコレステロール硫酸は、人工換気侵襲により放出される好中球エラスターゼを抑制し、酵素による侵襲から組織を防御していると推測された。 5.ラット好塩基球性白血病細胞(RBL cell)ヘロイコトリエンB4受容体(BLT)と血小板活性化因子受容体(platelet activating factor受容体:PAFR)の遺伝子を導入し、安定発現株を得た(RBL-BLT,RBL-PAFR)。それぞれの受容体を介する脱顆粒反応を測定しところ、RBL-BLT、RBL-PAFR共にLTB4及びPAFにより脱顆粒反応が認められた。EGTA処理により細胞外液中のカルシウムを除くと脱顆粒反応は抑制され、細胞内カルシウムの流入が必要であることが分かった。また百日咳毒素処理によりRBL-BLTではほぼ完全に脱顆粒反応が阻害されたが、RBL-PAFRでは中等度の阻害であり、BLTを介する脱顆粒反応はGiタンパク質を介し、PAFRを介する脱顆粒反応はGi及びGqタンパク質を介していると思われた。ホルボールエステル前処理によるconventional PKC(cPKC)の枯渇によってPAFRを介する脱顆粒反応のみが抑制され、cPKCの関与が示唆された。また、脱顆粒反応はチロシンキナーゼ阻害剤、MAPキナーゼ阻害剤処理により影響されなかったが、ホスファチジルイノシトール3キナーゼ(phosphatidylinositol-3 kiase:PI3K)阻害剤であるウオルトマニン及びLY294002、PKC阻害剤であるGF109203X処理により濃度依存的に抑制され、BLT及びPAFRを介する脱顆粒反応にはPKCとPI3Kが関与していることが示唆された。 6.RBL-BLT、RBL-PAFR共にLTB4及びPAFにより濃度依存的な細胞内カルシウム上昇反応が認められた。百日咳毒素処理によりLTB4依存性のカルシウム上昇は抑えられたが、PAF依存性の上昇は抑えられなかった。BLTを介する細胞内カルシウム上昇反応はGiタンパク質を介していると思われた。 7.RBL-BLT、RBL-PAFRにおけるホスファチジルイノシトール3リン酸(PIP3)産生を測定したところ、LTB4及びPAFにより一過性にPIP3が産生された。PIP3産生は百日咳毒素処理により強く抑制され、Giタンパク質を介していると思われた。また、EGTAやBAPTAといった細胞内外のカルシウムキレート剤の影響を受けなかったことから、細胞内カルシウム濃度上昇は必要ないことが示唆された。 以上、本論文は硫酸化脂質コレステロール硫酸が消化管上皮と気管支肺胞洗浄液中に存在することを示し、コレステロール硫酸の膵エラスターゼ及び好中球エラスターゼに対する阻害作用を明らかにした。また、ラット好塩基球性白血病細胞を用いて、LTB4及びPAFによる脱顆粒反応には、細胞内カルシウムの上昇に加え、PKCとPI3Kが必要であることを明らかにした。本研究は、リソソーム酵素の内因性阻害物質を明らかにし、さらに細胞内でのリソソーム放出の分子機構を明らかにすることで、白血球による初期炎症反応のメカニズムの解明に重要な貢献をなすと考え、学位の授与に値するものと考えられる。 | 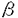 -hexosaminidaseの活性を人工基質を用いて測定した。様々なシグナル分子の阻害剤による阻害実験においては、各薬剤を20分間作用させた後、リガンド刺激を行った。
-hexosaminidaseの活性を人工基質を用いて測定した。様々なシグナル分子の阻害剤による阻害実験においては、各薬剤を20分間作用させた後、リガンド刺激を行った。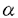 1-アンチトリプシンと複合体を形成し、不活化されることで活性が調節されている。しかし、両酵素に対する攻撃を受けることになる上皮組織内に存在する生体内防御物質は、これまで知られていなかった。消化管上皮組織において、硫酸化脂質コレステロール硫酸とスルファチドにより膵エラスターゼ酵素活性が抑制され、酵素による自己融解から粘膜を保護している可能性が考えられた。また、全身麻酔中の気管支肺胞洗浄液より、好中球エラスターゼとコレステロール硫酸が同時に検出され、コレステロール硫酸が好中球エラスターゼ酵素活性を用量依存的に抑制することが明らかになった。肺胞上皮由来の硫酸化脂質により酵素による侵襲が防御されている可能性が考えられた。また、ステロイド骨格をもつ他のステロイド硫酸は酵素活性に影響しないことから、コレステロール硫酸のもつ疎水性側鎖と、スルファチドとの共通構造である硫酸基が、酵素との複合体形成に必要であると考えられた。
1-アンチトリプシンと複合体を形成し、不活化されることで活性が調節されている。しかし、両酵素に対する攻撃を受けることになる上皮組織内に存在する生体内防御物質は、これまで知られていなかった。消化管上皮組織において、硫酸化脂質コレステロール硫酸とスルファチドにより膵エラスターゼ酵素活性が抑制され、酵素による自己融解から粘膜を保護している可能性が考えられた。また、全身麻酔中の気管支肺胞洗浄液より、好中球エラスターゼとコレステロール硫酸が同時に検出され、コレステロール硫酸が好中球エラスターゼ酵素活性を用量依存的に抑制することが明らかになった。肺胞上皮由来の硫酸化脂質により酵素による侵襲が防御されている可能性が考えられた。また、ステロイド骨格をもつ他のステロイド硫酸は酵素活性に影響しないことから、コレステロール硫酸のもつ疎水性側鎖と、スルファチドとの共通構造である硫酸基が、酵素との複合体形成に必要であると考えられた。