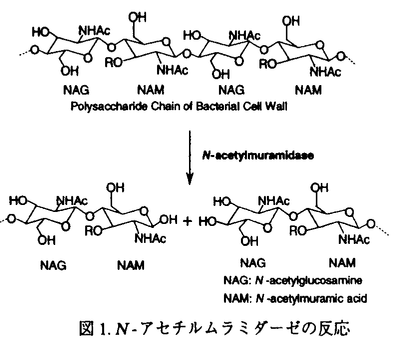
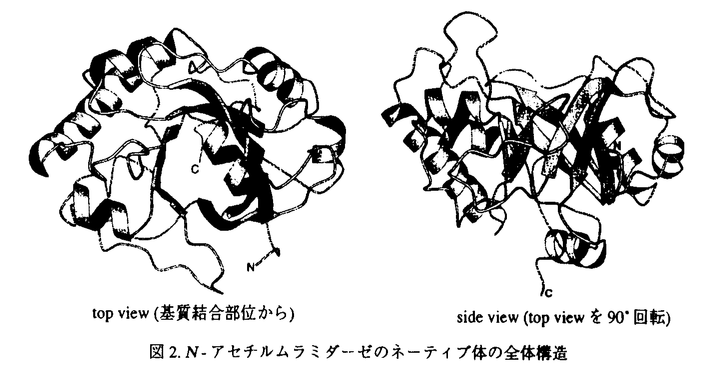
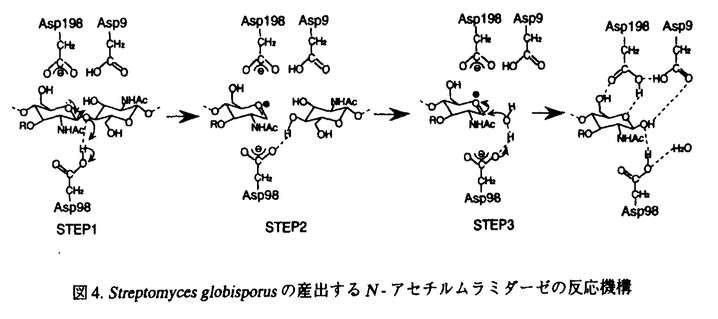
N-アセテルムラミダーゼは、グラム陽性菌の細胞壁多糖を形成しているN-アセチルムラミン酸(NAM)とN-アセチルグルコサミン(NAG)間の -1,4グリコシド結合を加水分解する酵素である。生体が細菌の感染から身を守るという重要な役割を果たしているため、このN-アセチルムラミダーゼは、多くの生物種に広く分布している。Streptomyces globisporusが産出するN-アセチルムラミダーゼは、ニワトリ卵白リゾチームが溶菌できない虫歯の原因菌Streptococcus mutansに作用するのに対して、ニワトリ卵白リゾチームが加水分解するキチン(ポリ-1,4-N-アセチルグルコサミン)は加水分解しない。論文での研究は、S.globisporusのN-アセチルムラミダーゼの三次元構造をX線結晶構造解析で明らかにし、酵素反応のメカニズムや基質認識機構を解明している。 -1,4グリコシド結合を加水分解する酵素である。生体が細菌の感染から身を守るという重要な役割を果たしているため、このN-アセチルムラミダーゼは、多くの生物種に広く分布している。Streptomyces globisporusが産出するN-アセチルムラミダーゼは、ニワトリ卵白リゾチームが溶菌できない虫歯の原因菌Streptococcus mutansに作用するのに対して、ニワトリ卵白リゾチームが加水分解するキチン(ポリ-1,4-N-アセチルグルコサミン)は加水分解しない。論文での研究は、S.globisporusのN-アセチルムラミダーゼの三次元構造をX線結晶構造解析で明らかにし、酵素反応のメカニズムや基質認識機構を解明している。 N-アセチルムラミダーゼは、16%ポリエチレングリコール8000を含む0.1M Tris-HCl緩衝液(pH8.6)を結晶化剤とするハンギングドロップ蒸気拡散法で、良質な結晶を再現性良く得られている。X線構造解析は本酵素とアミノ酸配列に約50%の相同性をもつStreptomyces erythraeusが産生するN-アセチルムラミダーゼを探索モデル分子とする分子置換法で行われている。2.05 分解能で三次元構造の精密化を行なった結果、結晶学的信頼度因子Rが0.186で三次元構造が決定された。アミノ酸残基数217から成る三次元構造は、図に示すように、8本の鎖が集まってバレル構造を作り、これが分子の中心にあってコアを形成し、その周りを6本の 分解能で三次元構造の精密化を行なった結果、結晶学的信頼度因子Rが0.186で三次元構造が決定された。アミノ酸残基数217から成る三次元構造は、図に示すように、8本の鎖が集まってバレル構造を作り、これが分子の中心にあってコアを形成し、その周りを6本の -ヘリックスが取り囲んでいる。また、8本の鎖のうちC末端側の鎖のみが逆平行に配置し、C末端に存在する-ヘリックスはバレルの下側(最初の鎖のN末端側)に蓋をするように配置している。全体としての分子の大きさは、約50×40×40である。 -ヘリックスが取り囲んでいる。また、8本の鎖のうちC末端側の鎖のみが逆平行に配置し、C末端に存在する-ヘリックスはバレルの下側(最初の鎖のN末端側)に蓋をするように配置している。全体としての分子の大きさは、約50×40×40である。  図.S.globisporusN-アセチルムラミダーゼのネーティブ体の全体構造 図.S.globisporusN-アセチルムラミダーゼのネーティブ体の全体構造 次に、本酵素の酵素反応の機構を解明するために、反応生成物の2-acetamido-2-deoxy-D-glucose--1,4-2-acetamido-2-deoxy-D-muramic acid(NAG-NAM)との複合体結晶を、基質特異性に関する知見を得るために、基質類似物2-amino-2-deoxy-D-glucose--1,4-2-amino-2-deoxy-D-glucose((GlcN)2)との複合体結晶を、それぞれソーキング法で調製した。X線回折強度データは120Kにおいて分解能1.8まで収集し、結晶学的な三次元構造の精密化を行った。得られた主な知見は以下の通りである。 NAG-NAM分子はシートで構成されているバレルのC末端側にあるクレフトに結合し、クレフト周辺に存在している酸性アミノ酸残基の側鎖とは水素結合で、芳香性アミノ酸残基の側鎖とはvan der Waals接触で相互作用していることを示した。NAG-NAM分子の結合様式から、NAMのO1酸素原子の部分で基質が加水分解されると考えた。このO1酸素原子には、芳香性のアミノ酸残基の側鎖に囲まれて疎水的な環境にあるAsp98の側鎖のカルボキシル基の酸素原子が、3.29の距離にある。また、O1酸素原子とAsp98の両方に水素結合している水分子が存在する。したがって、本酵素の酵素反応の機構として次のように考察した。まず、Asp98が酸触媒となり、基質のNAMとNAGの間の-1,4グリコシド結合中の酸素原子にプロトンを引き渡し、グリコシド結合の切断が起る。この時、反応中間体として生じるオキソカルベニウムイオンはAsp198の負電荷で安定化される。最後に,今度はAsp98が塩基触媒となり水分子を活性化し、その結果生じたOH-がO1酸素原子と隣接する炭素原子を求核攻撃し反応が終了する。この機構は、Streptomyces erythraeusが産生するN-アセチルムラミダーゼでも、これらのアミノ酸残基が一次配列と三次元構造上、保存されているという知見と合致する。さらに、触媒基と考えているAsp98が疎水的な環境にあることは、本酵素の至適pHが6.5であるという知見を説明する。 (GlcN)2との複合体でも、(GlcN)2分子はNAG-NAM分子と同様、本酵素のシートで構成されているバレルのC末端側にあるクレフトに結合していた。しかし、O1酸素原子が、(GlcN)2複合体では、触媒基のAsp98ではなく、Tyr62のO 原子と2.68の距離で水素結合している。また、反応中間体のオキソカルベニウムイオンで正電荷をもつO5酸素原子は、NAG-NAM複合体では、Asp198のO 原子と2.68の距離で水素結合している。また、反応中間体のオキソカルベニウムイオンで正電荷をもつO5酸素原子は、NAG-NAM複合体では、Asp198のO 原子が水素結合しているのに対して、(GlcN)2複合体では、Asp198のO原子とは5.83とかなり離れている。以上のことから、グルコサミンの多量体であるキトサンでは、本来の基質とは結合様式が異なり、酵素反応が進行しないものと考えられる。 原子が水素結合しているのに対して、(GlcN)2複合体では、Asp198のO原子とは5.83とかなり離れている。以上のことから、グルコサミンの多量体であるキトサンでは、本来の基質とは結合様式が異なり、酵素反応が進行しないものと考えられる。 このように、本論文は、これまでに見い出されているリゾチーム群とは基質特異性を異にするバクテリア由来のN-アセチルムラミダーゼの三次元構造を解明し、その構造と機能の相関を詳細に明らかにしている。よって、蛋白質の構造化学の進歩に寄与するところ大であり、博士(薬学)の学位を授与するに十分な内容を有すると認める。 |