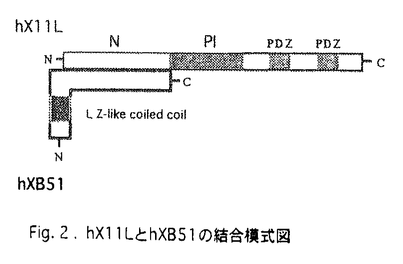
X11-like(X11L)は、アルツハイマー病(AD)の原因因子の1つとされている -アミロイド前駆体タンパク質(APP)の細胞質ドメインと相互作用するタンパク質としてYeast Two-hybrid法を用いてcDNAが単離された。X11Lは、分子中央部にPhosphotyrosine Interaction(PI)ドメインを、PIドメインのC-末端側に2つのPDZドメインを持ち、すでに報告されていたX11とアミノ酸配列上で53.5%の相同性を示すLIN10遺伝子ファミリーに所属する分子である。X11LはAPPに由来する-アミロイドの生成を制御することが明らかにされてきたが、その機能は不明な点が多い。しかしながら、(1)X11LのPiドメインのN-末端側の配列が、APPとの相互作用に影響を与えること、(2)X11LとX11は、PIドメインのN-末端側の配列で類似性が乏しく(アミノ酸配列上で18.2%の相同性)、N-末端側の配列が両分子の機能特異性を規定している可能性があることが考えられた。そこで、X11Lのアミノ末端側の配列に結合し、X11Lの機能を修飾する分子の探索と機能解析を行った。 -アミロイド前駆体タンパク質(APP)の細胞質ドメインと相互作用するタンパク質としてYeast Two-hybrid法を用いてcDNAが単離された。X11Lは、分子中央部にPhosphotyrosine Interaction(PI)ドメインを、PIドメインのC-末端側に2つのPDZドメインを持ち、すでに報告されていたX11とアミノ酸配列上で53.5%の相同性を示すLIN10遺伝子ファミリーに所属する分子である。X11LはAPPに由来する-アミロイドの生成を制御することが明らかにされてきたが、その機能は不明な点が多い。しかしながら、(1)X11LのPiドメインのN-末端側の配列が、APPとの相互作用に影響を与えること、(2)X11LとX11は、PIドメインのN-末端側の配列で類似性が乏しく(アミノ酸配列上で18.2%の相同性)、N-末端側の配列が両分子の機能特異性を規定している可能性があることが考えられた。そこで、X11Lのアミノ末端側の配列に結合し、X11Lの機能を修飾する分子の探索と機能解析を行った。 まず、ヒトX11L(hX11L)の126-555アミノ酸をコードするcDNAをbaitとして、ヒト脳cDNAライブラリーよりYeast Two-hybrid法を用いてスクリーニングを行った。得たクローンの中、#51と名付けたクローンは273アミノ酸からなる新規タンパク質をコードする完全長cDNAであった。このタンパク質をX11L binding protein clone #51(XB51)と名付けた。ヒトXB51(hXB51)cDNAをプローブにマウス完全長cDNA(mXB51)も単離した。mXB51は、分子中央部の32アミノ酸残基が欠失している以外はhXB51と75.5%の相同性を示した。XB51は分子のN-末端側(amino acid number17-55 of hXB51)にロイシンジッパー構造を持つ。 Northern blot hybridizationによる解析で、mXB51は脳特異的な発現を示し、hXB51は脳の他、心臓、骨格筋に強く発現が見られた。マウスプローブとヒトメンブレン、ヒトプローブとマウスメンブレンを用いたcross-probe hybridizationにおいても同様の結果が得られたので、アイソフォームが存在するのではなく、マウスとヒトでは発現様式が異なる事を明らかにした。hX11LとFLAG-hXB51の各ドメインコンストラクトをCOS7細胞に共発現させ、それぞれの特異抗体を用いた共役免疫沈降法で、両タンパク質の結合ドメインを同定した。その結果、XB51は、ロイシンジッパー構造を含まないC-末端側の配列でX11LのPIドメインよりN-末端側の領域と相互作用することを明らかにした。 XB51をCOS7細胞に安定的に発現させた場合、XB51は10mM CHAPS buffer不溶性画分に回収され、蛍光抗体染色により核周辺のER/cis-Golgiに局在することが明らかになった。しかしながら、X11Lを共発現させると10mM CHAPS buffer可溶性画分に回収され細胞質に分布するようになった。XB51は、X11L不在下ではER/cis-Golgiにアンカーリングされているが、X11Lと複合体を形成する事で、細胞内分布を変えることを示した。また、vitroにおける結合実験から、XB51はX11LとAPPの結合を非競合的、容量依存的に阻害することを明らかにした。 神経細胞におけるXB51の存在様式を検討する目的で、マウス脳およびヒトneuroblastoma SH-SY5Y細胞中のXB51を特異抗体を用いたWestern blot法で解析したところ、XB51タンパク質の他に約80kDaの高分子量タンパク質を検出した。脳および細胞の抽出液を6M塩酸グアニジン処理した後にWestern blot法を行ったところXB51だけを検出したことから、この約80kDaのタンパク質はXB51と未同定のタンパク質がSDS-耐性の複合体を形成しているものであると考えられた。XB51を安定的に発現するCOS7細胞を35S-メチオニンで代謝標識し、その抽出液とSH-SY5Y細胞の抽出液をインキュベートし、電気泳動後オートラジオグラフィーで解析したところ、約80kDaのタンパク質は出現した。しかしながら、放射標識されたXB51を含む抽出液を非神経細胞であるHEK293細胞の抽出液とインキュベートした場合は、約80kDaのタンパク質は検出出来なかった。この結果は、XB51と安定な複合体を形成しているタンパク質は神経特異的な分子であることを明らかにした。 XB51と複合体を形成している神経特異的タンパク質を同定する目的で、SH-SY5Y細胞を35S-メチオニンで代謝標識し、その抽出液から、XB51抗体を用いて複合体を回収し、6M塩酸グアニジン処理後SDS電気泳動で分離し、オートラジオグラフィーで解析したところ、XB51以外に55kDa,60kDaのタンパク質を検出した。これらのタンパク質をneuron-specific XB51binding protein with molecular weight 55kDa and 60kDa(NSX55/NSX60)と名付けた。XB51がNSXsに結合するドメインを決定するために、XB51のドメインコンストラクトを作製し、各コンストラクトを発現するCOS7細胞を35S-メチオニンで代謝標識し、その抽出液とSH-SY5Y細胞の抽出液をインキュベートし、電気泳動後オートラジオグラフィーで解析した。その結果、アミノ末端側のロイシンジッバー構造を欠失したXB51はNSXsと複合体を形成出来ないことを明らかにした。XB51とNSXsは互いにロイシンジッバーを介して、複合体を形成している事が示唆された。 神経細胞ではXB51はNSXsと安定な複合体を形成していることから、この複合体形成にX11Lがどのように関与しているのかを明らかにする目的で、SH-SY5Y細胞を35S-メチオニンで代謝標識し、その細胞抽出液にX11Lを加えてインキュベートし、XB51/NSXs複合体をXB51抗体を用いて回収後、オートラジオグラフィーで定量した。その結果、X11Lの容量依存的にXB51/NSXs複合体は減少したが、減少率は約35%に留まった。この結果、X11Lは非競合的にXB51/NSXs複合体形成を阻害するが、X11LとXB51の結合はXB51とNSXsの結合よりも弱いことが示唆された。 脳組織におけるXB51の分布を明らかにする目的で、成体マウス脳の連続切片を作製し、免疫染色を行った。その結果、観察した脳内の神経細胞の細胞質および神経突起にXB51が発現していることを明らかにした。発現は大脳皮質、海馬の神経細胞で強い傾向が認められた。 XB51とアルツハイマー病との関連を調べる目的で、家族性アルツハイマー病(FAD),弧発性アルツハイマー病(SAD),健常老人脳におけるXB51/NSXsの存在様式を生化学的に解析した。その結果、FAD,SAD共に、アルツハイマー病患者の脳では、XB51/NSXs複合体が10mM CHAPSbuffer不溶性画分に大部分が回収された。健常人脳では、大部分のXB51/NSXs複合体は10mMCHAPS buffer可溶性画分に回収された。著しい量的な変化は認められなかった。また、APPやX111Lの存在様式に量的・質的変化はなかった。このことは、XB51/NSXs複合体の不溶化が何らかの神経機能障害を引き起こし、神経変性を引き起こす原因になっている可能性を示唆する。一連の研究はAPP-X11L-XB51-NSXsというprotein Interaction mappingを解明し、神経変性の分子機構を解明する上でその重要性を示した。 以上、本研究において、新規X11L結合タンパク質XB51のcDNAを単離し、その構造機能を解析すると共に、X11Lを中心としたprotein Interaction mappingを解明し、アルツハイマー病患者ではXB51/NSXs複合体の存在様式に違いがあることを明らかにした。これらの新規に発見した事実を多く含んでいる知見は、アルツハイマー病の発病分子機構を解明する上で重要な知見を与えると共に、原因因子とされるAPPが細胞内でどのような位置にいるのかに関する知見を与え、protein Interactionの異常が発病の原因となりうる新たな概念を提示したことで、博士(薬学)の授与に値するものと判断した。 |