プララミノーゲンアクナベーター(PA)には組織型PA(tissue-type PA:tPA)とウロキナーゼ型PA(urokinase-type PA:uPA)とがあり、PAは前駆体であるプラスミノーゲンをプラスミンに変換し、そして基質特異性の低いプラスミンが多くの細胞外マトリックス(extracellular matix:ECM)を分解するとされている。中枢神経系においてPA-プラスミン系は細胞が移動する際に活性化し、その周辺部のECMを溶解しながら進路の形成に寄与することが明らかにされている。しかし動物が成熟して活発な神経活動を行うときのシナプス可塑性や、更には精緻な構造体である神経回路が形成されるときのシナプス形成に対するプラスミンの影響については不明な点が多い。近年、PA-プラスミン系は癲癇、痙攣、興奮毒性時などで海馬を含めた脳内で亢進することが報告され、その亢進時の役割について精力的に研究がすすめられている。本研究ではPA-プラスミン系の亢進を想定し、脳の重要な機能である学習と記憶を司る海馬において、プラスミンのシナプス可塑性およびシナプス形成に対する影響を検討した。 1.シナプス可塑性に対するプラスミン前処置の影響 長期の薬物適用に適した培養海馬切片を実験に用いた。8日齢のWistar系ラットより厚さ300 mの海馬切片を作成し、血清含有培地に浸したテフロン膜上で培養した。培養10-15日後、100nMプラスミンを含む無血清培地で6時間の前処置を施した。その後、Schaffer側枝刺激によってCA1野放線層に誘発される場の興奮性後シナプス電位(field excitatory postsynaptic potential:fEPSP)の傾きをシナプス伝達の指標として測定し、機能的シナプス可塑性の一例である長期増強(LTP:long-term potentiation)を高頻度刺激で誘導した。また免疫蛍光法を用いてECMの発現を検討した。 mの海馬切片を作成し、血清含有培地に浸したテフロン膜上で培養した。培養10-15日後、100nMプラスミンを含む無血清培地で6時間の前処置を施した。その後、Schaffer側枝刺激によってCA1野放線層に誘発される場の興奮性後シナプス電位(field excitatory postsynaptic potential:fEPSP)の傾きをシナプス伝達の指標として測定し、機能的シナプス可塑性の一例である長期増強(LTP:long-term potentiation)を高頻度刺激で誘導した。また免疫蛍光法を用いてECMの発現を検討した。 プラスミンを含有する培地は測定前に洗浄した。コントロールの切片群では高頻度刺激により持続的なLTPが認められたが、プラスミン前処置群ではLTPは誘導されたもののその後期相の維持が障害された(図1A)。プラスミンを洗浄し更に24-48時間培養した切片群では、通常のLTPが観察された。また高頻度刺激なしのシナプス伝達に対して、プラスミン前処置は影響を及ぼさなかった(図1B)。以上の結果より、プラスミン前処置が可逆的にLTPの維持を障害し、そしてこの障害は単なるシナプス応答の減衰によるものではないことが示唆された。  図1 プラスミン前処置のLTP(A)およびシナプス伝達(B)に及ぼす影響。○:コントロールの切片群、●:プラスミン100nMで6時間前処置した切片群、■:プラスミンを洗浄し更に24-48時間培養した切片群。 図1 プラスミン前処置のLTP(A)およびシナプス伝達(B)に及ぼす影響。○:コントロールの切片群、●:プラスミン100nMで6時間前処置した切片群、■:プラスミンを洗浄し更に24-48時間培養した切片群。 LTPの維持にプラスミシによって分解されたECMが必須ではないかと考え、ECMの主要な構成成分であるラミニン-1、フィブロネクチン、IV型コラーゲンの発現を免疫蛍光法を用いて検討した。ラミニンは海馬錐体細胞層および海馬と嗅内皮質内の血管壁に強く発現していた(図2A)。プラスミン前処置は全領域においてラミニンの発現を減少させ(図2B)、プラスミンを洗浄し更に24-48時間培養した切片群では再びラミニンの強い発現が認やられた(図2C)。フィブロネクチンおよびコラーゲンも海馬内に発現が認められたものの、いずれもプラスミン前処置による影響を受けなかった。  図2 プラスミン前処置のラミニンに及ぼす影響。A.コントロールの切片、B.プラスミン100nMで6時間前処置した切片、C.プラスミンを洗浄し更に24-48時間培養した切片。bar=500m。 図2 プラスミン前処置のラミニンに及ぼす影響。A.コントロールの切片、B.プラスミン100nMで6時間前処置した切片、C.プラスミンを洗浄し更に24-48時間培養した切片。bar=500m。 LTPの障害がプラスミン前処置によるラミニンの分解に起因すると考えられた。そこでラミニンの分解を阻害することにより、プラスミン前処置によるLTPの減弱効果が抑制されるかを検討した。ラミニンに選択的に結合しプラスミンによるラミニンの分解を抑制すると予期された抗ラミニン抗体をその阻害剤として使用した。抗ラミニン抗体で処置した後、プラスミン前処置した切片群では、プラスミン前処置によるラミニンの分解が阻害された(図3A)。次にこの切片でのLTPを検討したところ、高頻度刺激により通常のLTPが認められた(図3B)。以上の結果よりプラスミンによるラミニンの分解がLTP障害の主要因であり、ラミニン以外の基質が分解されたとしてもその寄与は小さいことが示唆された。 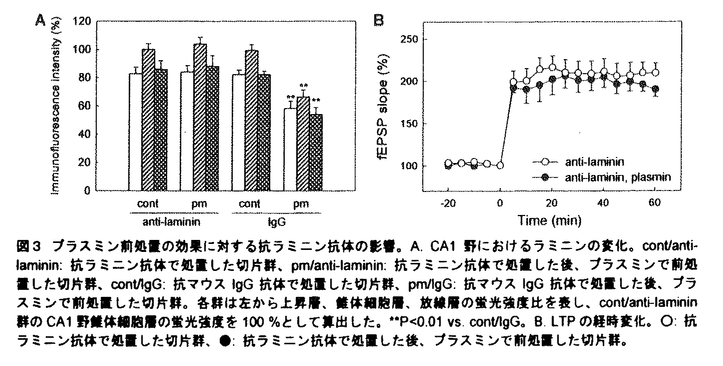 図3 プラスミン前処置の効果に対する抗ラミニン抗体の影響。A.CA1野におけるラミニンの変化。cont/anti-laminin:抗ラミニン抗体で処置した切片群、pm/anti-laminin:抗ラミニン抗体で処置した後、プラスミンで前処置した切片群、cont/IgG:抗マウスIgG抗体で処置した切片群、pm/IgG:抗マウスIgG抗体で処置した後、プラスミンで前処置した切片群。各群は左から上昇層、錐体細胞層、放線層の蛍光強度比を表し、cont/anti-laminin群のCA1野錐体細胞層の蛍光強度を100%として算出した。**P<0.01vs.cont/IgG。B.LTPの経時変化。○:抗ラミニン抗体で処置した切片群、●:抗ラミニン抗体で処置した後、プラスミンで前処置した切片群。2.シナプス形成に対するプラスミン慢性処置の影響 図3 プラスミン前処置の効果に対する抗ラミニン抗体の影響。A.CA1野におけるラミニンの変化。cont/anti-laminin:抗ラミニン抗体で処置した切片群、pm/anti-laminin:抗ラミニン抗体で処置した後、プラスミンで前処置した切片群、cont/IgG:抗マウスIgG抗体で処置した切片群、pm/IgG:抗マウスIgG抗体で処置した後、プラスミンで前処置した切片群。各群は左から上昇層、錐体細胞層、放線層の蛍光強度比を表し、cont/anti-laminin群のCA1野錐体細胞層の蛍光強度を100%として算出した。**P<0.01vs.cont/IgG。B.LTPの経時変化。○:抗ラミニン抗体で処置した切片群、●:抗ラミニン抗体で処置した後、プラスミンで前処置した切片群。2.シナプス形成に対するプラスミン慢性処置の影響 培養期間中の培養海馬切片では切片作製時の障害が修復され、再び神経回路が構築されるため、切片レベルでのシナプス形成を評価するのが可能である。この培養海馬切片を100nMプラスミンを含む血清含有培地で培養することによりプラスミンの慢性処置をおこなった。Schaffer側枝刺激によってCA1野錐体細胞層に誘発される集合スパイク(PS)の最大振幅をシナプス形成の指標として測定した。また胎生18日齢Wistar系ラットより単離した海馬神経細胞を用いてプラスミン慢性処置の神経突起の形態に及ぼす影響を検討した。 コントロールの切片群と比べて、プラスミン慢性処置群では集合スパイクが有意に減少した(図4A)。プラスミン慢性処置のECMに対する影響を検討したところ、ラミニンとフィブロネクチンは分解されたものの、コラーゲンに対しては影響が認められなかった(図4B)。 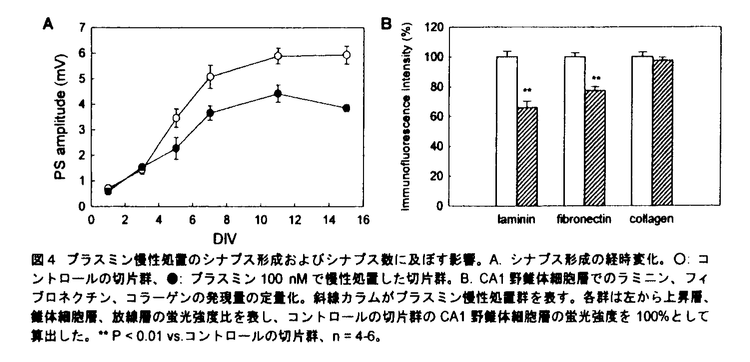 図4 プラスミン慢性処置のシナプス形成およびシナプス数に及ぼす影響。A.シナプス形成の経時変化。○:コントロールの切片群、●:プラスミン100nMで慢性処置した切片群。B.CA1野錐体細胞層でのラミニン、フィブロネクチン、コラーゲンの発現量の定量化。斜線カラムがプラスミン慢性処置群を表す。各群は左から上昇層、錘体細胞層、放線層の蛍光強度比を表し、コントロールの切片群のCA1野錐体細胞層の蛍光強度を100%として算出した。**P<0.01vs.コントロールの切片群、n=4-6。 図4 プラスミン慢性処置のシナプス形成およびシナプス数に及ぼす影響。A.シナプス形成の経時変化。○:コントロールの切片群、●:プラスミン100nMで慢性処置した切片群。B.CA1野錐体細胞層でのラミニン、フィブロネクチン、コラーゲンの発現量の定量化。斜線カラムがプラスミン慢性処置群を表す。各群は左から上昇層、錘体細胞層、放線層の蛍光強度比を表し、コントロールの切片群のCA1野錐体細胞層の蛍光強度を100%として算出した。**P<0.01vs.コントロールの切片群、n=4-6。 プロテアーゼによるECMの分解は神経細胞の形態に深く関与するとされているため、プラスミン慢性処置が神経突起の形態に影響を及ぼしていると考えられた。そこで海馬神経細胞を用いてプラスミン慢性処置の最長突起長、最長突起の分岐数、シナプス数に対する影響を検討した。ラミニンコーティングは最長突起長を有意に増加させた。プラスミン慢性処置は培養4日目において最長突起長を抑制する傾向を示したもの、有意には抑制しなかった。一方、分岐数に対してラミニンコーティング自体は影響を与えなかったものの、ラミニンコーティング上でのプラスミン慢性処置は有意に分岐数を減少させた。FM1-43を用いてシナプス数を検討したところ、ラミニンはシナプス数を有意に増加させ、プラスミン慢性処置はポリリジンのみのコート上では影響を及ぼさなかったものの、ラミニンによって増加したシナプス数を有意に減少させた(図5A)。以上の結果よりプラスミン慢性処置が機能的シナプス数を減少させ、シナプス形成を阻害する可能性が示唆された。更にラミニンの細胞内情報伝達を検討するため、mitogen actitivated protein kinase(MAPK)を活性化するMAPK kinaseの阻害剤であるU0126およびphosphatidylinositol 3-kinaseの阻害剤であるLY294002を添加したところ、U0126はラミニンによるシナプス数の増加を抑制し、LY294002は影響を及ぼさなかった。 次にプラスミン慢性処置がMAPKのリン酸化に影響を及ぼしているかを検討した。ラミニンは細胞膜上の接着蛋白質であるインテグリンと相互作用をし、さらにインテグリンの細胞内ドメインに結合するfocal adhesion kinase(FAK)が細胞内情報伝達を司ると考えられている。最初にFAKの発現を検討したところ、FAKの発現量がラミニンによって増加し、プラスミン慢性処置によって減少していた(図5B)。またプラスミン慢性処置はMAPKの発現量に影響を及ぼさなかったが、ラミニンによるMAPKのリン酸化を抑制した。 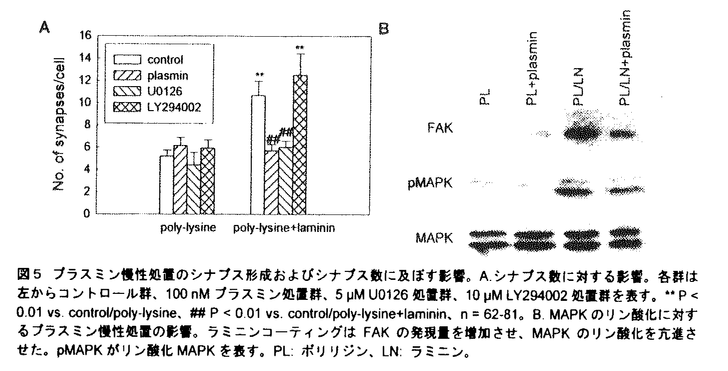 図5 プラスミン慢性処置のシナプス形成およびシナプス数に及ぼす影響。A.シナプス数に対する影響。各群は左からコントロール群、100nMプラスミン処置群、5M U0126処置群、10M LY294002処置群を表す。**P<0.01vs.control/poly-lysine、##P<0.01vs.control/poly-lysine+laminin、n=62-81。B.MAPKのリン酸化に対するプラスミン慢性処置の影響。ラミニンコーティングはFAKの発現量を増加させ、MAPKのリン酸化を亢進させた。pMAPKがリン酸化MAPKを表す。PL:ポリリジン、LN:ラミニン。 図5 プラスミン慢性処置のシナプス形成およびシナプス数に及ぼす影響。A.シナプス数に対する影響。各群は左からコントロール群、100nMプラスミン処置群、5M U0126処置群、10M LY294002処置群を表す。**P<0.01vs.control/poly-lysine、##P<0.01vs.control/poly-lysine+laminin、n=62-81。B.MAPKのリン酸化に対するプラスミン慢性処置の影響。ラミニンコーティングはFAKの発現量を増加させ、MAPKのリン酸化を亢進させた。pMAPKがリン酸化MAPKを表す。PL:ポリリジン、LN:ラミニン。 本研究はシナプス可塑性およびシナプス形成に対するプラスミンの影響を検討し、以下のことを明らかにした。 1.培養海馬切片においてプラスミン前処置はラミニンの分解を介してLTPを調節する。 2.プラスミン慢性処置はラミニン、フィブロネクチンを分解し、また集合スパイクを減少させる。 3.培養海馬神経細胞においてプラスミン慢性処置はラミニンコート上でシナプス数を減少させる。 本研究により、中枢神経系においてPA-プラスミン系が亢進した場合にはラミニンの分解を介したシナプス可塑性の減弱およびラミニンとフィブロネクチンの分解を介したシナプス形成の阻害が引き起こされる可能性が示唆された。多くの神経疾患でPA-プラスミン系は亢進することが報告されており、今後、PA-プラスミン系によるECMの分解と神経疾患との関連を明らかにすることが望まれる。 |