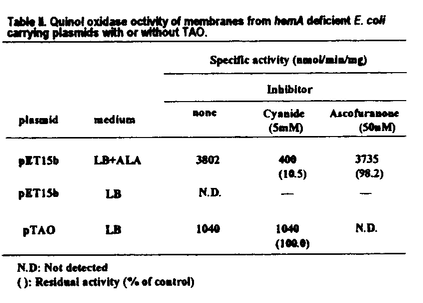
アフリカ型トリパノソーマ(Trypanosoma brucei)は、ツェツェバエによって媒介されるアフリカ睡眠病の病原体となる原虫であり、またT.bruceiは、ヒトの寄生虫感染症としてばかりでなく、動物、特に家畜の感染がアフリカの食糧問題におけるタンパク源欠乏の中心的な問題となっている。アフリカ型トリパノソーマはその表面抗原を変異させ、宿主の免疫システムから回避する事がよく知られているが、そのため有効なワクチンの開発は困難をきわめている。この様な状況において、抗トリパノソーマ薬による化学療法は非常に重要な位置を占めており、宿主であるヒトと大きく異なった性質を持つミトコンドリアのエネルギー代謝系は極めて有望な標的と考えられる。 これまでの研究でT.bruceiのミトコンドリアはその生活環において機能を大きく変化させることが明らかになっている。すなわち、ベクターであるツェツェバエ中のプロサイクリック型ではATP合成は酸化的リン酸化によって行われているが、宿主である哺乳類の血液中に生息する血流型でのATP合成はトリパノソーマに特有なオルガネラであるグリコソーム中で行われる解糖系の反応に依存している。この中で血流型のミトコンドリアはグリコソームで生じる還元力を解消するグリセロール-3-リン酸(G-3-P)酸化系を形成しており、シトクロム電子伝達系は存在しない。このG-3-P酸化系の末端酸化酵素であるTrypanosome alternative oxidase(TAO)はシアン耐性で、哺乳類に存在するシアン感受性のいわゆるシトクロムc酸化酵素とは全く異なった構造を持っている。このTAOは宿主であるヒトなど哺乳類には存在しない点から、化学療法の格好の標的と考えられる。実際にTAOを標的とする研究が欧米を中心に精力的に進められており、我が国でも原虫ミトコンドリアのG-3-P依存の酸素消費およびユビキノール依存の酸素消費をnMオーダーで阻害する新規抗トリパノソーマ剤アスコフラノンが見い出されている。この薬剤は感染マウスにおいて、グリセロールとの併用投与によって投与後120分以内にトリパノソーマを血液中から完全に消滅させ、その有効性はin vivoでも確かめられている。最近の遺伝子レベルの研究から、TAOと植物に存在するシアン耐性末端酸化酵素との共通性が明らかになってきたが、酵素が不安定で単離精製が困難である事から生化学的解析は進んでいない。そのため、これら薬剤の作用機構をはじめとして多くの問題が未解決の状況にある。本研究はこの問題を解決する目的で大腸菌における組み替えTAO発現系の構築を試み、さらにTAOを標的とした新しい薬剤のスクリーニング系の確立をめざして行われたものである。 まず、第一段階としてアフリカ型トリパノソーマ原虫から精製したmRNAより、RT-PCRにて増幅したTAO遺伝子(CDNA)を発現ベクターpET15bに挿入しで、TAO発現ベクター(pTAO)を作成した。pTAOを導入した大腸菌BL21(DE3)pLysSにはCBB染色にて分子量約37kDaの新たなペプチドのバンドを認め、このペプチドは植物シアン耐性末端酸化酵素に対するモノクローナル抗体によって認識された。また、その局在は膜画分であったが、キノール酸化活性を調べたところ、pTAOを導入した大腸菌ではシアン存在下でも全く阻害されなかった。さらにpTAOを導入した大腸菌の増殖はシアンに耐性で、TAOの性質を獲得しており、大腸菌において活性のある組み替えTAOの発現系が確立された。 次に、大腸菌のヘム合成系欠損変異株を宿主とした発現系の構築を行った。その理由は通常、大腸菌は呼吸鎖中にTAOと同じキノール酸化活性を触媒する大腸菌由来の二つの末端酸化酵素、シトクロムboおよびbd複合体を持っており、以後の種々の解析を不明瞭なものにするからである。そこで、大腸菌末端酸化酵素の構成成分であるヘムの合成能を欠損したhemA欠損株を用いた発現系の確立を試みた。hemA欠損株は呼吸鎖中の末端酸化酵素および複合体IIの活性が失われることが知られているが、培養液中へのアミノレブリン酸(ALA)の添加により人為的にヘムの合成を制御することができる。BL21(DE3)にP1形質導入を行い作成したヘム欠損株FN102にコントロールベクターを導入したFN102/pET15bの膜画分の活性はシアンに感受性でアスコフラノンに耐性であった。またこの株をNAを添加せず、培養した膜画分のキノール酸化活性は検出限度以下であった。一方、pTAOを導入した株FN102/pTAOではその膜画分はアミノレブリン酸無添加の条件下でもキノール酸化活性を示し、その活性はシアン耐性でアスコフラノンに感受性であった。この様に組み換えTAOを唯一の末端酸化酵素として持つ大腸菌の発現系が確立された。さらに、この組み換えTAOとトリパノソーマ原虫から調製したミトコンドリアのキノール酸化活性の酵素学的な性質について比較したところ、電子供与体であるユビキノールに対するKm値、阻害剤であるアスコフラノンに対するKi値ともに同程度の値を示し、本来の性質を保持した組み換えタンパク質の発現系である事が確認された。 また、FN102/pET15bとFN102/pTAOのLB培地中での好気的増殖を比較検討したところFN102/pET15bの倍加時間約4時間に対してFN102/pTAOの倍加時間約1時間となり、組み換えTAOの存在により増殖が早くなることが分かった。このFN102/pTAOの増殖はシアンに耐性であり、これはヘム欠損株の液体培地中での好気的増殖は組み換えTAOによって相補されていることを示していた。したがってFN102/pTAOは、TAOの阻害を好気的増殖の阻害として検出できることを意味しており、実際にFN102/pTAOをTAO阻害剤であるアスコフラノンおよびサリチルヒドロキサム酸を添加したLB培地中で好気的に培養したところ、アスコフラノン3nM〜10 M、サリチルヒドロキサム酸0.3M〜1mMにおいて濃度依存的に増殖が阻害された。つまりFN102/pTAOはTAOの阻害の程度を増殖の阻害として検出できることが明確になり、抗トリパノソーマ剤の新しいスクリーニング系が確立された。 M、サリチルヒドロキサム酸0.3M〜1mMにおいて濃度依存的に増殖が阻害された。つまりFN102/pTAOはTAOの阻害の程度を増殖の阻害として検出できることが明確になり、抗トリパノソーマ剤の新しいスクリーニング系が確立された。 以上、本研究ではヘム欠損株FN102を発現宿主とした組み換えTAOの大量発現系を構築し、原虫TAOと同様の性質を保持した組み換えTAOのみに由来するキノール酸化活性を持つ大腸菌膜画分を大量に調製する条件を確立することに成功した。さらにこの系を用いて、組み換えTAOの阻害の程度を増殖の阻害として検出する、抗トリパノソーマ剤の新しいスクリーニング系を確立した。この成果は、今まで生化学的、タンパク質化学的な性質が全く判っていなかったTAOの解析に極めて大きな進展を与えるとともに、TAOを標的とした抗トリパノソーマ剤のスクリーニング系の確立は新規抗トリパノソーマ薬の開発に大いに貢献すると考えられる。よって、博士(薬学)の学位に値すると判定した。 |