哺乳類中枢神経系においては、神経伝達物質や神経修飾物質が、相互に関連し合いながら神経活動を制御している。脳の機能を理解する上でこれらの物質間の修飾作用やクロストークは重要であり、これまでにも、様々な物質間の組み合わせで、形態学的立場からの同一細胞内共存やシナプス形成、生理学あるいは薬理学的立場からの機能的関連性が報告されている。本研究では、神経は飾物質としての神経ペプチドの作用の一例として、海馬におけるムスカリン性アセチルコリン受容体(mAChR)機能に及ぼすソマトスタチンの修飾作用について検討した。 海馬は、記憶、学習の成立に重要な役割を果しているが、この脳機能が障害される典型的な状態がアルツハイマー病である。この疾患では、海馬を中心として、特徴のある病理形態学的所見が見られると同時に、種々の神経化学的変化も現われる。この中で、アセチルコリン作動性神経の障害が最も顕著であり、古くから、アセチルコリンの合成酵素や分解酵素の活性、受容体結合能の低下が認められている。一方、神経ペプチドについてはソマトスタチンの異常が注目されている。実験的にも、アセチルコリン系やソマトスタチン系の機能低下が記憶、学習能力の障害を引き起こすことが知られており、海馬における記憶、学習の機能にはこの2つの物質系が関与していると考えられる。形態学的には、内側中隔核からのコリン作動性神経が、海馬のソマトスタチン含有神経細胞に直接入力することが認められている。このような事実から、海馬におけるアセチルコリン系とソマトスタチン系の機能的関連性の存在が推定され、in vitroでの受容体機能という観点から研究を進めることとした。具体的には、ラット海馬のmAChRの結合能及びその活性化後の細胞内反応に及ぼすソマトスタチンの影響を調べた。更に、そのメカニズムに関して、細胞内カルシウムイオン濃度([Ca2+])に対するこのペプチドの作用を検討した。 ウィスター系雄性ラットの海馬P2分画を用いて、mAChRの受容体結合実験を行なった。mAChR アゴニストの結合部位を観察するために、標識したアゴニストである3H-oxotremorine-M-acetate(oxo-M)結合の飽和実験を行なった。ソマトスタチンとして[D-trp8] somatostatin を反応溶液に加えた。その結果、[D-trp8]somatostatin は3H-oxo-M 結合の最大結合数を変化させずに親和性を低下させることが認められた。 この時、[D-trp8]somatostatin の作用は10-18Mから10-5Mの範囲で濃度依存性であった。また、mAChRアンタゴニストである3H-N-methyl-scopolamine(NMS)の阻害実験からoxotremorine結合部位を観察し、別の観点からmAChR アゴニスト結合をとらえた。oxotremorine結合部位は、通常高親和性と低親和性の2つの結合部位に分かれているが、[D-trp8] somatostatin添加により、高親和性結合部位が消失し低親和性結合部位のみの単一結合部位となった(図1)。一方、mAChR アンタゴニストとして3H-QNBと3H-NMS結合の飽和実験を行なった結果、mAChRアンタゴニスト結合部位は[D-trp8]somatostatinにより変化を受けないことがわかった。これらの成績から、ソマトスタチンは mAChR の受容体蛋白質以降の細胞内情報伝達系に作用し、mAChRアゴニスト結合部位の親和性を低下させることによりその機能状態を変化させているものと考えられた。 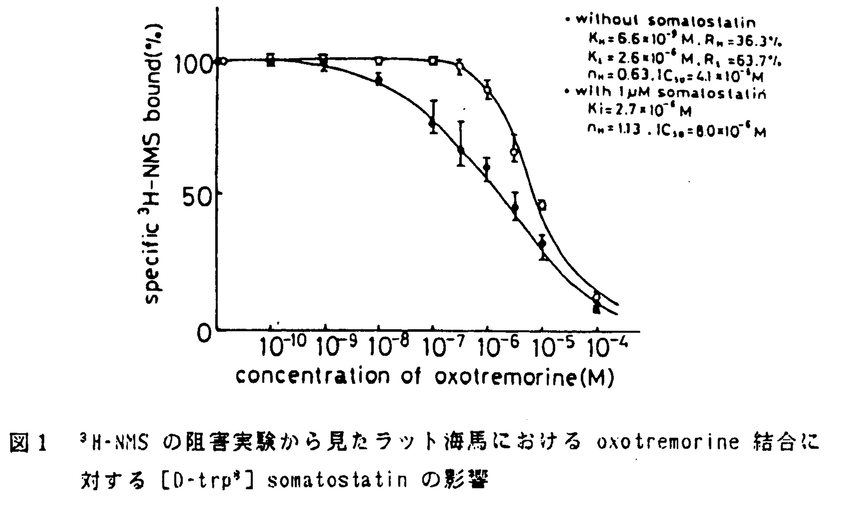 図1 3H-NMSの阻害実験から見たラット海馬におけるoxotremorine結合に対する[D-trp8]somatostatinの影響 図1 3H-NMSの阻害実験から見たラット海馬におけるoxotremorine結合に対する[D-trp8]somatostatinの影響 脳の部位による相違としては、上記のような mAChR アゴニスト結合部位に対するソマトスタチンの作用は、海馬や大脳皮質では認められたが、橋-延髄では見られなかった。海馬と大脳皮質は、総mAChRの80%がM1受容体であり、橋-延髄はその90%がM2受容体である。M1とM2受容体は、脳内分布や細胞内情報伝達機構に相違のあることが認められている。すなわち、M1受容体の活性化はイノシトールリン脂質(PI)代謝の亢進を引き起こし、M2受容体のそれはアデニル酸シクラーゼ活性を抑制する。このM1とM2受容体の細胞内情報伝達機構の差が、ソマトスタチンの部位特異的な作用発現の原因ではないかと考えられた。 そこで、M1受容体に対するソマトスタチンの影響を調べるために、ラット海馬目のスライスを用いてPI代謝の測定実験を行った。スライスに3H-myo-inositolを取り込ませた後、mAChR アゴニストであるカルバコール、ソマトスタチンあるいはその両者による刺激を与え、生成される3H-inositol-1-phos-Phate の量を測定した。カルバコールは、従来からの報告通り、海馬におけるPI 代謝を亢進させた。[D-trp8] somatostatin は、それ自身PI代謝を変化させなかったが、 カルバコールによるPI代謝の亢進を更に増強させた。この[D-trp8]somatostatinの作用は、5×10-10Mから5×10-5Mの間で濃度依存性を示した。このようなソマトスタチンによる、アゴニスト結合の親和性の低下と受容体に連関する細胞内応答の増強は、このペプチドがmAChRの機能を活性化していることを意味するものと推測された。 次に、以上のような海馬におけるソマトスタチンの mAChR に及ぼす影響のメカニズムを調べる目的で、このペプチドの[Ca2+]iに対する作用を検討した。これは、 PI代謝がCa2+依存性の系であることから、 ソマトスタチンがmAChRに連関するPI代謝を増強させるという実験成績が[Ca2+]iの変動によって説明可能ではないかと考えられたからである。実験としては、胎生18日齢のラットから海馬を取り出し、約2週間培養した神経細胞を用いて、[Ca2+]iを測定した。細胞にfura-2を取り込ませた後、340nmと360nmの励起光を当て、発生する光をoptical fiberでとらえた。[D-trp8]somatostatin を培養神経細胞に灌流投与すると、急峻な立ち上がりの一過性の[Ca2+]i上昇が見られた(図2)。この作用は、濃度依存性であった。細胞外液をCa2+-freeにすると、[D-trp8]somatostatin の作用は完全に抑えられた。また、N型Ca2+チャンネル拮抗薬である  -conotoxin GVIA で細胞を前処理すると、[D-trp8] somatostatin の効果は完全に抑制された。これらのことから、海馬においてソマトスタチンはN型Ca2+チャンネルを通して細胞外液からの Ca2+流入を引き起こしていることが示唆された。 -conotoxin GVIA で細胞を前処理すると、[D-trp8] somatostatin の効果は完全に抑制された。これらのことから、海馬においてソマトスタチンはN型Ca2+チャンネルを通して細胞外液からの Ca2+流入を引き起こしていることが示唆された。 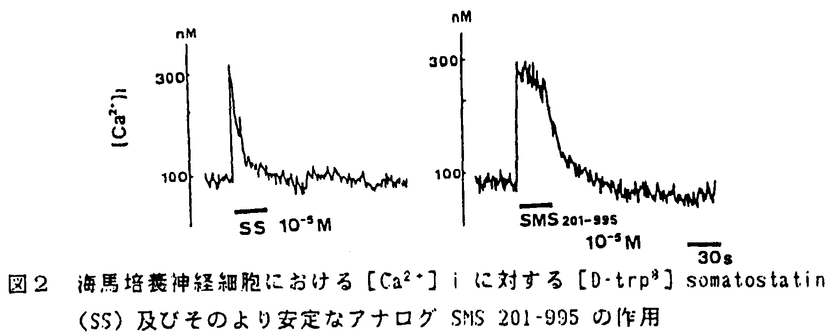 図2 海馬培養神経細胞における[Ca2+]iに対する[D-trp8]somatostatin(SS)及びそのより安定なアナログ SMS 201〜995の作用 図2 海馬培養神経細胞における[Ca2+]iに対する[D-trp8]somatostatin(SS)及びそのより安定なアナログ SMS 201〜995の作用 従来から、ソマトスタチンは抑制性の神経ペプチドとして知られ、種々の組織でホルモン分泌、膜電位、アデニル酸シクラーゼ活性、[Ca2+]i に対し抑制性に作用することが知られている。本実験において、海馬でソマトスタチンが[Ca2+]i上昇という興奮方向への作用を有することを見い出したことは注目に値する。また、ソマトスタチンが[Ca2+]iを上昇させることを前提とすれば、このペプチドが mAChR 結合の親和性を低下させることや、この受容体に連関するPI代謝を増強させることが容易に納得され、この細胞内応答がmAChR機能の修飾において重要であると考えられた。 |