| 【緒言】 骨粗鬆症の本質は骨量の減少とそれにもとずく骨の脆弱性の亢進であり、本症は骨折の危険率を増加させる。その中で、閉経後骨粗鬆症は女性ホルモンであるエストロゲンの低下に対する適応不全がその一因として考えられている。臨床的にもエストロゲン投与は骨量減少を予防し、骨粗鬆症治療に有効性を示す。しかしながら、エストロゲンの骨に対する作用機序は未だ明らかとなっていない。特に、エストロゲン受容体(ER)の機能や発現様式には、不明な点が数多く残されている。骨におけるERの存在や、エストロゲンの直接作用を検出する試みは従来から行われていたが、否定的なものが多かった。しかし、1988年骨芽細胞に、1991年破骨細胞に蛋白レベルならびに遺伝子レベルでERの存在が証明され、エストロゲンが骨においてその受容体を介して直接的な作用を持つ可能性が示唆された。 本研究では、免疫組織化学的手法を用い3種の骨芽細胞様細胞におけるERの発現を検討したところ、ERを発現している細胞としていない細胞のあることが観察された。そこで、このERの発現が細胞周期に依存するという仮説を設け、細胞周期とERの発現との関連につき、免疫組織化学的手法、ウェスタンブロット解析ならびにノーザンブロット解析により検討した。さらに、細胞周期を同調させた骨芽細胞様細胞を用い、エストロゲンの細胞増殖に及ぼす作用についても検討を加えた。 【実験方法】 細胞培養:MC3T3-E1細胞(マウス骨芽細胞株),HOS TE85細胞(ヒト骨肉腫細胞株),MCF-7細胞(ヒト乳癌細胞株)は、60mg/mlのkanamycin monosulfateならびに10%ウシ胎児血清を含有する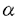 -MEM培地で37℃、5%CO2の条件下にて培養した。新生児マウスの頭蓋骨由来の骨芽細胞は、Raizsらの方法に従い0.1%collagenaseおよび0.2%dispaseを含有するphosphate buffer salineで分離精製し、上述の環境下で培養した。 -MEM培地で37℃、5%CO2の条件下にて培養した。新生児マウスの頭蓋骨由来の骨芽細胞は、Raizsらの方法に従い0.1%collagenaseおよび0.2%dispaseを含有するphosphate buffer salineで分離精製し、上述の環境下で培養した。 ERの免疫染色:ERの検出は、細胞を中性4% paraformaldehydeで固定後、ERのDNA-binding domainを特異的に認識するマウス抗ERモノクロナール抗体(MA1-310)ならびに核内ERを選択的に認識するマウス抗ERモノクロナール抗体(NCL-ER-LH1)を用い、streptavidin-biotin peroxidase酵素法ならびにstreptavidin-biotin phycoerythrin(PE)蛍光法により行なった。 二重蛍光免疫染色:HOS TE85細胞を用い、ERおよび細胞周期のS期(DNA合成期)のマーカーであるBromodeoxyuridine(BrdU)の二重蛍光免疫染色を行なった。BrdU(10-5M)を30分間細胞に取り込ませた後に中性4% paraformaldehydeで細胞を固定した。ERの蛍光染色は抗ER抗体を用いたstreptavidin-biotin PE蛍光法により行なった。BrdUの蛍光発色はfluorescein-5-isothiocyanate-マウス抗BrdUモノクロナール抗体を使用した。二重蛍光染色は上述の2種類の抗ER抗体で行った。 S期開始時期への同調培養:HOS TE85細胞がsubconfluentになった時点で、thymidine(2.5×10-3M)およびhydroxyurea(10-3M)を順次処理し、S期開始時期に同調させた。S期開始後、経時的に細胞数ならびにBrdUの取り込みを測定し、細胞周期の進行をモニターした。 BrdUの取り込み実験:細胞にBrdU(10-5M)を30分間取り込ませた後、中性4% paraformaldehydeで固定した。BrdUの免疫染色はマウス抗BrdUモノクロナール抗体を使用したstreptavidin-biotin peroxidase酵素法により行なった。BrdUを取り込んだ細胞の評価は、異なる4視野で100個づつ計400個の細胞を観察し、抗BrdU抗体で染色された細胞の割合を%で表示した。 ウエスタンブロット解析:細胞周期の各相におけるHOS TE85細胞から核分画を抽出し、ウエスタンブロット法によりナイロンフィルターを作製した。フィルター上のER蛋白は、抗ER抗体(NCL-ER-LH1)を用いたstreptavidin-biotin alkaline phosphatase酵素法により行なった。 ノーザンブロット解析:細胞周期の各相におけるHOS TE85細胞からtotal RNAを抽出した。ノーザンブロット法により作製したナイロンフィルターは、32P-dCTPで標識したラットER cDNAプローブによるhybridizationに供し、引き続きオートラジオグラフィーを行った。 同調培養したHOS TE85細胞における17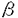 -estradiolの増殖に対する作用:S期開始時に同調したHOS TE85細胞に17-estradiolを添加し、[3H]thymidineの取り込み、細胞数の変化ならびにBrdUを取り込いだ細胞数を経時的に計測した。 -estradiolの増殖に対する作用:S期開始時に同調したHOS TE85細胞に17-estradiolを添加し、[3H]thymidineの取り込み、細胞数の変化ならびにBrdUを取り込いだ細胞数を経時的に計測した。 [3H]thymidineの取り込み実験:同調したHOS TE85細胞に17-estradiolならびに[3H]thymidineを添加し、24時間後にDNAに取り込まれた[3H]thymidineの放射活性を計測した。 【結果】 ERの免疫染色:3種の骨芽細胞様細胞において、ERが細胞質ならびに核内に検出された。しかし、そのERの発現に関しては細胞間の差が大きく、多様性が見られた。また、3種の骨芽細胞様細胞への17-estradiol(10-gM)の添加はERの発現に著明な影響を示さなかった。なお陽性対照として用いたMCF-7細胞において、17-estradiolの添加はERの発現を減少した。 ノーザンブロット解析によるER mRNAの検出:MC3T3-E1細胞およびHOS TE85細胞において、ノーザンブロット解析により6.5kbおよび6.2kbのバンドが検出された。これらのバンドは報告されているER mRNAの大きさと一致した。17-estradiolの添加はER mRNAの発現にも著明な変動をもたらさなかった。 ERとBrdUの二重蛍光免疫染色:HOS TE85細胞において、ERはBrdUが取り込まれたS期の細胞に強く発現する傾向が観察された。これは、ERの異なる領域を認識する2種類の抗ER抗体で、同様の結果であった。 S期開始時期への同調培養:S期開始時期に同調したHOS TE85細胞の細胞数とBrdUの取り込みの推移から、この細胞の細胞周期の進行は以下の様に推定された。即ち、S期開始より1.5時間後では95%以上、3時間後では約90%、6時間後では約70%の細胞がS期に位置する。12時間経過時では全体の85%以上がG2期(分裂準備期)およびM期(分裂期)に移行する。18時間後ではほとんどの細胞がG2期およびM期を終了し、21ならびに24時間経過時ではG1期(合成準備期)および次周期のS期の細胞が混在する。 細胞周期の各相におけるERの発現:上述の同調培養したHOS TE85細胞を用い、ERの発現を検討した。免疫染色によりERはS期初期に細胞質で検出されはじめ、S期の中期以後には細胞質および核に分布し、その後、ERはG1期に減少することが観察された。ところが、次の細胞周期のS期に再び発現してくることが認められた。ウエスタンブロット解析においても、同様の結果が得られた。さらに、ノーザンブロット解析によりER mRNAはS期初期に強く発現し、免疫染色されるER蛋白に先がけて発現することが示された。ER mRNAは、その後時間経過とともに減少し、次の周期のS期には再び発現してくることが確認された。 同調培養したHOS TE85細胞における17-estradiolの増殖に対する作用:17-estradiolは、S期開始後24時間の[3H]thymidineの取り込みを増加し、15ならびに24時間に細胞数を有意に増加した。さらに、17-estradiolによりS期開始後21時間にBrdUを取り込んだ細胞は有意に増加した。 【考察】 骨芽細胞で観察されたER発現の多様性は細胞周期に依存することが示された。これは、骨芽細胞のエストロゲンに対する応答性が、細胞周期の各相に依存する可能性を示唆している。細胞周期に依存したERの発現は酵母やヒト乳癌細胞(G1期に発現)においても報告されているが、発現時期については本研究で見だされた骨芽細胞のものとは異なっている。これは各組織のERの発現様式に違いがあることを表しており、各組織におけるエストロゲン応答性やエストロゲンの作用機作の相違を示唆している。また、本研究により、エストロゲンが骨芽細胞の細胞周期の進行を促進することが判明した。エストロゲンはERが多く発現しているS期およびG2/M期に作用し、細胞周期の進行および細胞の機能を調節している可能性が考えられる。近年、エストロゲンが骨形成に作用することが報告されているが、本研究よりERは、成長期、骨折からの治癒過程ならびに癌化等の骨の細胞が急激に増殖する時期において重要な役割を担うことが示唆される。 |