本研究は、単細胞性緑藻クラミドモナスの細胞表層カーボニックアンヒドラーゼ(CA)のタンパク質および遺伝子の単離と、CO2濃度条件に対応した遺伝子発現機構を解明する目的で行われた。論文は5章よりなる。 第1章では、高いCO2濃度で培養した細胞からのCAタンパク質の単離とその性質について述べている。クラミドモナスを連続光照射下、高いCO2条件(5%CO2)で培養し、アフィニティクロマトグラフィーと、DEAEイオン交換HPLCを用いてCAを単離した。その結果、180mlの細胞より0.2mgのCAが得られた。HPLCビークのリテンションタイムの違いから、単離したCAは、これまで研究されてきた低CO2条件下で発現するCAとは異なることがわかった。そこで、低CO2条件で発現するCAをCA1とし、高CO2条件で培養した細胞に存在するCAをCA2と命名した。CA2は、SDS-ゲル電気泳動の結果から、CA1と同様に大小2つのサブユニットからなり、その分子量は大サブユニットが38kDa、小サブユニットが4.2kDaであった。DTTで還元処理すると活性は消失することから、CA2もCA1と同様にサブユニット間がジスルフィド結合によって会合していること、また、フコースに特異的なレクチンと反応することから、CA2はCA1と同様にフコースを含む糖タンパクであることが明らかになった。 第2章では、CA2がCAH2遺伝産物であることを確認した実験について述べている。CA2の大小サブユニットのアミノ末端部位のアミノ酸配列を決定した。CAH1、CAH2の両遺伝子配列を比較した結果、大サブユニットのアミノ末端から15、16残基目、また小サブユニットでは1、3、4、12、13残基目のアミノ酸残基が異なっていた。そこで、CA2についてこれらのアミノ酸残基を解析した。小サブユニットでは、1残基目でグルタミン酸残基、3残基目でチロシン残基等が見いだされ、残りすべての残基においてもCAH2に対応するアミノ酸残基が確認できた。このことより、高いCO2条件で培養した細胞から得られたCA(CA2)はCAH2遺伝子産物であることが明らかとなった。すなわち、クラミドモナスの細胞表層には2つのCAアイソザイムが存在することが結論づけられた。 第3章ではCAH1遺伝子、CAH2遺伝子の発現に及ぼす酢酸の影響について検討している。CAH1、CAH2の2つの遺伝子のmRNAに特異的な合成オリゴヌクレオチドプローブを作成し、これを用いてノーザンハイブリダイゼーションを行い、その発現量を調べた。独立栄養下で培養した細胞では、低CO2条件に移すとCAH1 mRNAは1時間目から現れ、2時間目で最大となり、4時間目で定常的なレベルに達するのに対し、CAH2 mRNAは高CO2条件で発現し、低CO2条件に移すと1時間目には消失した。酢酸を与えた従属栄養下で培養した細胞では、低いCO2条件に移すと、CAH1 mRNAは4時間目に弱い発現が見られたが、6時間目にはほぼ消失した。一方、CAH2のmRNAは低いCO2条件に移したにもかかわらず高いCO2条件と同じ程度の発現が確認された。 第4章ではCAH1遺伝子、CAH2遺伝子の発現に及ぼす気体の影響について検討している。N2、O2ガスについて調べたところ、CAH1、CAH2のmRNAの発現量にはほとんど影響は見られなかった。これらのことにより、それぞれのmRNAの発現にはCO2濃度が大きく関与しており、N2、O2の影響はCO2濃度の影響に比べほとんどないか、あっても非常に小さいことが明らかになった。さらに、CO2濃度による発現への影響を詳細に調べた。CAH1 mRNAは、0%から0.3%とCO2濃度が高まるにつれて、その転写量が減少するに対し、CAH2 mRNAは逆に増加した。最大の発現量はCAH1 mRNAとCAH2 mRNAのそれぞれで大きく違うことから、それぞれ最大に発現する量を100%として、各CO2濃度での発現の相対量を調べた。その結果、CAH1、 CAH2のそれぞれのmRNAの発現量はCO2濃度0.1%を中心にミラーイメージとなることが明らかになった。以上の結果から、CA1は光合成の行われているとき、特にCO2濃度の低い条件で発現しているのに対し、CA2は光合成に関係なく、CA1の発現が抑えられているときに発現していることが明らかになった。 第5章ではCAH1遺伝子、CAH2遺伝子の5’上流域の比較を行った結果について述べている。調べた全ての条件において、転写レベルで発現調節されることから、各遺伝子の発現調節に関与すると思われる5’上流域の塩基配列を決定し、解析した結果CAH1とCAH2の5’上流域に共通配列が見いだされた。そこでCAと同じように複数の遺伝子からなるRuBisCO、tubulinの5’上流域を比較したところ、RuBisCO、tubulinにそれぞれ共通な配列が存在することが明らかとなった。なお、CAH1、CAH2は、遺伝子はともに1コピーであり、それぞれ細胞あたりの存在比にしてmRNAは60-70:1、タンパク質は100:1であった。このことより、CA1とCA2の最大発現量の差は、主として転写量の差によると考えられた。 以上のように本論文では、クラミドモナスのカーボニックアンヒドラーゼの生化学的特徴と遺伝子発現について、炭酸ガス濃度の高低という環境条件の変化を実験系に取り入れ、巧みに解析を行い、2種類のカーボニックアンヒドラーゼが存在するという新しい事実を明らかにした。環境の変化に適応した遺伝子の発現機構に関してはまだ未知の部分が多く、今後も多くの研究が成されるものと考えられるが、本研究は機構の全容を解明する上で大きな貢献を成すものであり、その成果は高く評価できる。第1章、第2章に関しては、既に公表されており、第3章に関しては印刷中である。また、第4章、第5章に関しては近く公表予定である。いずれも共著論文であるが、本論文提出者が研究の主要部分を担当したものであることを確認した。ここに審査員一同は、論文提出者立木光は東京大学博士(学術)の学位を授与するに値するものと認める。 | 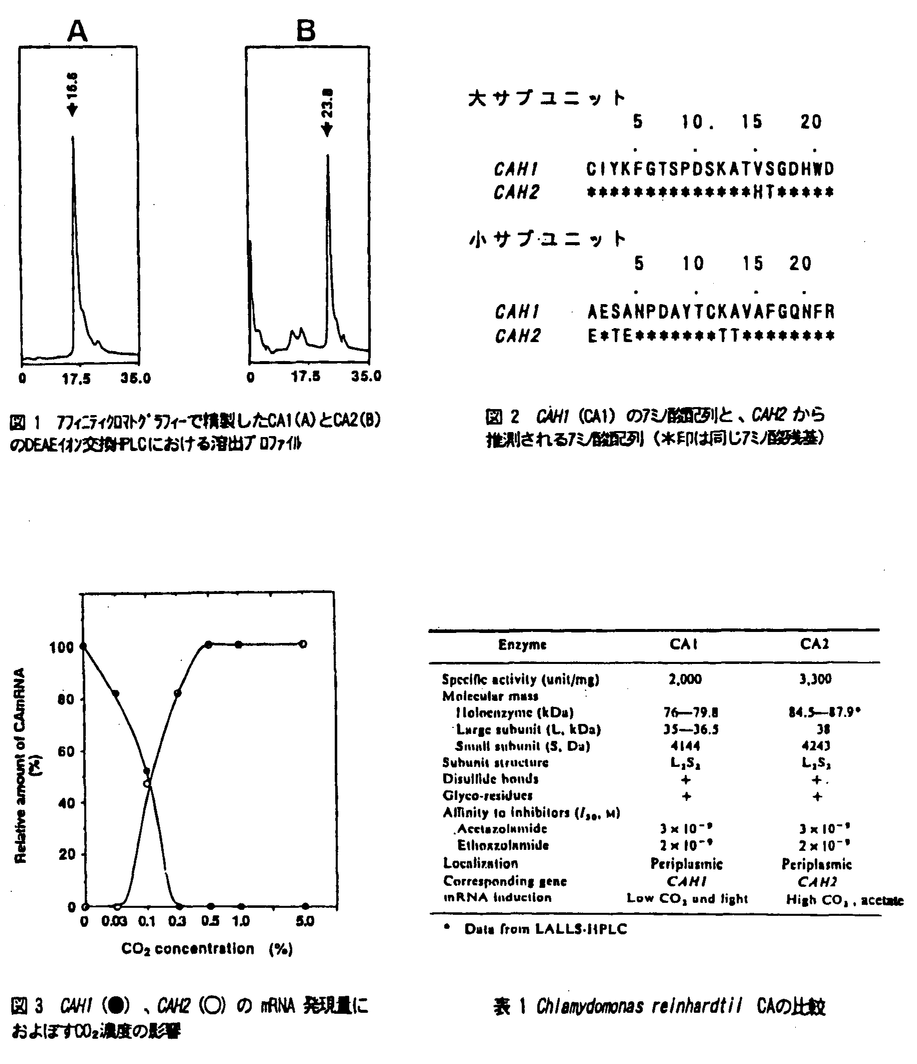
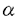 1,
1,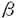 1,
1,