| 内容要旨 | | 酵母のキラー現象は,ある種の酵母がタンパク質性のキラー因子を分泌して他の酵母の生育を阻害する酵母間の拮抗現象である。これまでに多種の酵母が多様な性質のキラー因子を分泌することが示されてきたが,キラー因子及びキラー遺伝子の構造あるいはその作用機作について分子レベルの解析が進んでいるキラー現象は,数多く報告されたキラー現象のごく一部に過ぎない。本研究では,日本の伝統的発酵食品中の耐塩性キラー酵母に着目し,それらの分布及び特性を調べるとともに,新規キラー酵母の検索を行った。その結果得られた新規キラー酵母Pichia farinosa KK1株に焦点を絞り,キラー因子及びキラー遺伝子の構造解析を行った。 1.発酵食品中の耐塩性キラー酵母の解析 日本各地から伝統的発酵食品である味噌,醤油,漬物を収集し,食塩存在下でキラー活性を示すキラー酵母を分離した。51点の試料のうち28点から56株のキラー酵母が分離された。そのうち53株はDebaryomyces hanseniiであり,のこり3株はCandida naeodendra,Hansenula anomala,Pichia farinosaと同定された。C.naeodendraとP.farinosaはこれまでキラー酵母としての報告は無く,新規なキラー酵母である。漬物,塩漬け野菜の試料からは76%という高率で主としてD.hanseniiに属するキラー酵母が分離された。味噌,醤油等の試料からは18点中,3点からキラー酵母が分離されたにとどまった。分離したキラー酵母について,既にキラー酵母としての特性で分類されている標準キラー酵母(K1-K10)に対する食塩存在下及び非存在下におけるキラー活性を調べた。その結果,53株のD.hanseniiのキラースペクトルは多様であり,またその多様性は食塩存在下でより増加する傾向が見られた。他の3株も食塩存在下でより多くの酵母に対してキラー活性を示す特徴を持つことが明らかになった。 2.Pichia farinosa KK1株の生産するキラー因子,SMKトキシンの解析 味噌麹から分離した新規キラー酵母P.farinosa KK1株の生産するキラー因子は,食塩存在下でより強いキラー活性を示すことから,Salt Mediated KillerよりSMKトキシンと命名し,解析を行った。まず,キラー活性を高感度に検出できるアッセイ法として,マイクロタイタープレートを用いる希釈法を確立した。次いでこのアッセイ法を用いて培養条件の検討を行い,比活性の高い培養濾液を得た。SMKトキシンの安定性を調べた結果,pH4以下で活性は安定に保たれるが,pH4.5以上では安定性が減少し,pH6では完全に失活すること,またキラー活性は温度の上昇と共に減少し,50℃以上では30分間で完全に失活することが明らかとなった。またSMKトキシンはNaCl及びKClの濃度の増加に伴ってキラー活性が上昇することも明らかとなった。 SMKトキシンの精製は,硫安沈澱,SP-トヨパールによるイオン交換クロマトグラフィー,Butyl-トヨパールによる疎水性クロマトグラフィーにより行った。活性画分の酸性条件でのポリアクリルアミドゲル電気泳動及び低分子ペプチドの分離が可能なトリシンSDS-ポリアクリルアミドゲル電気泳動(SDS-PAGE)による解析の結果,SMKトキシンはトリシンSDS-PAGE上で4kDaと8kDaを示す と と のサブユニットからなる2量体であることが明らかになった。両サブユニット間にはジスルフィド結合は無く,酸性(pH4.5以下)から中性へのpH変化で両サブユニットは容易に解離すること,解離と同時にサブユニットは凝集して沈澱しサブユニットは溶液中に残ることが明らかとなった。一方,酸性溶液中のSMKトキシンのサブユニット間の結合は強く,1.5M食塩存在下あるいは6M尿素存在下でも2量体として挙動した。両サブユニットのアミノ酸組成分析,アミノ酸配列分析の結果から,サブユニットは疎水性アミノ酸に富む約60アミノ酸残基からなるペプチドであること,サブユニットは約75アミノ酸残基からなるペプチドであることが推定された。 のサブユニットからなる2量体であることが明らかになった。両サブユニット間にはジスルフィド結合は無く,酸性(pH4.5以下)から中性へのpH変化で両サブユニットは容易に解離すること,解離と同時にサブユニットは凝集して沈澱しサブユニットは溶液中に残ることが明らかとなった。一方,酸性溶液中のSMKトキシンのサブユニット間の結合は強く,1.5M食塩存在下あるいは6M尿素存在下でも2量体として挙動した。両サブユニットのアミノ酸組成分析,アミノ酸配列分析の結果から,サブユニットは疎水性アミノ酸に富む約60アミノ酸残基からなるペプチドであること,サブユニットは約75アミノ酸残基からなるペプチドであることが推定された。 3.SMKトキシンをコードする遺伝子SMK1の解析 サブユニットから得られたアミノ酸配列を元にプライマーを作製し,P.farinosa KK1株のゲノムDNAを鋳型にしてPCR分析を行った。サブユニットのリジルエンドペプチダーゼ断片の配列をコードする170-bpのPCR断片をプローブに用いて,ゲノムDNAのEcoR I完全消化物を gt11に組み込んだライブラリーのスクリーニングを行った。得られた5.5-kbのEcoR I断片に含まれる1.4-kbのHindIII断片の塩基配列分析を行った結果,222アミノ酸をコードする666-bpのオープンリーディングフレーム(ORF)か見いだされた。サブユニットのN末端配列はORFのGly-146からの配列と完全に一致した。またサブユニットのリジルエンドペプチダーゼ断片の配列から,サブユニットのC末端はORFにコードされる最後のアミノ酸残基Asp-222であることが推定された。このORF上のMet-1からAla-18の配列は典型的なシグナルペプチドの特徴を有していた。続くTrp-19からの配列はサブユニットのN末端アミノ酸配列と一致し,両サブユニットが同一の遺伝子上にコードされていることが判明した。サブユニットのC末端ペプチドのアミノ酸配列分析の結果,C末端アミノ酸はVal-81と同定された。したがって,サブユニットはそれぞれ63及び77アミノ酸残基からなるペプチドであることが明らかになった。ORF上でVal-81に続く2残基はLys-Argの塩基性アミノ酸であり,これは酵母及び高等真核生物で見られるプロセシング酵素Kex2エンドプロテアーゼの認識配列である。サブユニットのC末端がVal-81であることは,SMKトキシンの生成においても,ORFにコードされる前駆体から,Kex2類似酵素による2残基の塩基性アミノ酸のC末端側の切断,続いてKex1類似酵素によるC末端の塩基性アミノ酸の除去,というプロセシングが行われたことを示唆している(図1)。 gt11に組み込んだライブラリーのスクリーニングを行った。得られた5.5-kbのEcoR I断片に含まれる1.4-kbのHindIII断片の塩基配列分析を行った結果,222アミノ酸をコードする666-bpのオープンリーディングフレーム(ORF)か見いだされた。サブユニットのN末端配列はORFのGly-146からの配列と完全に一致した。またサブユニットのリジルエンドペプチダーゼ断片の配列から,サブユニットのC末端はORFにコードされる最後のアミノ酸残基Asp-222であることが推定された。このORF上のMet-1からAla-18の配列は典型的なシグナルペプチドの特徴を有していた。続くTrp-19からの配列はサブユニットのN末端アミノ酸配列と一致し,両サブユニットが同一の遺伝子上にコードされていることが判明した。サブユニットのC末端ペプチドのアミノ酸配列分析の結果,C末端アミノ酸はVal-81と同定された。したがって,サブユニットはそれぞれ63及び77アミノ酸残基からなるペプチドであることが明らかになった。ORF上でVal-81に続く2残基はLys-Argの塩基性アミノ酸であり,これは酵母及び高等真核生物で見られるプロセシング酵素Kex2エンドプロテアーゼの認識配列である。サブユニットのC末端がVal-81であることは,SMKトキシンの生成においても,ORFにコードされる前駆体から,Kex2類似酵素による2残基の塩基性アミノ酸のC末端側の切断,続いてKex1類似酵素によるC末端の塩基性アミノ酸の除去,というプロセシングが行われたことを示唆している(図1)。 ペプチドと名付けたプロセシングにより切り放されるとの間のSer-84からArg-145の配列には,ORF中で唯一の推定上のN結合型糖鎖の結合位置であるAsn-127が存在する。SHKトキシンとSaccharomyces cerevisiaeのK1トキシンは1次構造上の類似性は無いにもかかわらず,その前駆体構造,疎水性度,プロセシングの様式等において非常に類似していることが明らかになった。 ペプチドと名付けたプロセシングにより切り放されるとの間のSer-84からArg-145の配列には,ORF中で唯一の推定上のN結合型糖鎖の結合位置であるAsn-127が存在する。SHKトキシンとSaccharomyces cerevisiaeのK1トキシンは1次構造上の類似性は無いにもかかわらず,その前駆体構造,疎水性度,プロセシングの様式等において非常に類似していることが明らかになった。 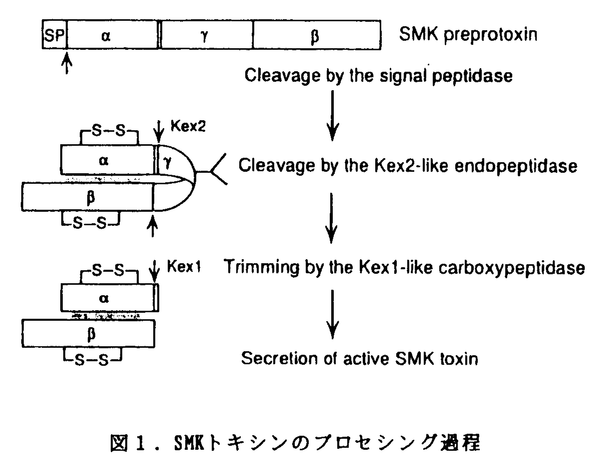 図1.SMKトキシンのプロセシング過程 図1.SMKトキシンのプロセシング過程 ノーザンブロット解析およびプライマー伸長法による解析の結果,SMK1のmRNAは長さが約1-kbであり,5’末端としてnt-218と-311の2カ所が同定された。nt-218の50-bp上流には典型的なTATA boxが存在していた。 染色体電気泳動の結果,P.farinosa KK1の染色体は少なくとも5本のバンドに分離され,SMK1はその最大のバンドとハイブリッド形成したことからSMK1が染色体上に存在することが確認された。 4.抗サブユニット抗体に反応する糖タンパク質Gp26の解析 P.farinosa KK1株を6%食塩を含むYPD培地(1%酵母エキス,2%ポリペプトン,2%グルコース)で培養した培養濾液中には,ウェスタンブロットにより抗サブユニット抗体と反応する約26kDaのタンパク質(Gp26)が存在することが明らかになった。培養条件の検討を行った結果,Gp26の分泌には0.5M以上の食塩を含むYPD培地が必要であり,食塩濃度2-2.5MでGp26の分泌が最大になった。食塩の有無にかかわらずYM培地(0.3%酵母エキス,0.3%麦芽エキス,0.5%ポリペプトン,1%グルコース)あるいは10%の味噌抽出液を含むYM培地ではGp26は分泌されなかった。Gp26の精製は硫安沈澱,SP-トヨパールによるイオン交換クロマトグラフィー,Biofine HIC-PHによる疎水性クロマトグラフィーにより行った。ナタマメの-mannosidase及びEndo--N-acetylglucosaminidase(Endo H)による酵素消化を行った結果,SDS-PAGE上で-mannosidaseでは500程度,Endo Hでは2000程度分子量の減少が見られたことから,Gp26が糖タンパク質であることが確認された。アミノ酸配列分析の結果,Gp26のN末端配列はSMKトキシンのサブユニットのN末端配列と一致した。また,Gp26のリジルエンドペプチダーゼ断片の配列は,サブユニット及びORF上に存在するペプチドの配列と一致した。以上の結果からGp26はSMKトキシンのプロトキシンであることが明らかになった。ノーザンブロット解析の結果,SMK1のmRNAは食塩非存在下でも定常的に転写されていることが示され,食塩により誘導されるGp26の分泌は転写以降の翻訳あるいは分泌経路の過程で制御されていると推定された。 |
| 審査要旨 | | 本論文は,高濃度食塩存在下で生育する耐塩性酵母のキラー現象に注目し,新規キラー酵母の分離およびそのキラー因子の構造の解明を目的として実施した研究結果をまとめたもので,本文4章および緒言と総括よりなる。 第I章では,発酵食品からの耐塩性キラー酵母の分離同定と,分離されたキラー酵母の特性解析について論じている。本論文の著者は,日本各地から伝統的発酵食品である味噌,醤油,漬物を収集し,食塩存在下でキラー活性を示すキラー酵母を分離した。分離した56株のキラー酵母のうち,53株はDebaryomyces hanseniiであり,のこり3株はHansenula anomala,Cangida naeodendua,Pichia farinosaと同定された。C.naeodendraとP.farinosaはこれまでキラー酵母としての報告は無く,新規なキラー酵母である。食塩存在下及び非存在下におけるキラー活性を調べた結果,53株のD.hanseniiのキラースペクトルは多様であり,他の3株も食塩存在下においてより多くの酵母に対してキラー活性を示すという特徴を持つことが明らかになった。 第II章は,Pichia farinosa KK1株の生産するキラー因子,SMKトキシンについて論じたものである。P.farinosa KK1株の生産するキラー因子は,食塩存在下でより強いキラー活性を示すことから,SMKトキシン(Salt Mediated killer toxin)と命名した。SMKトキシンの活性はpH4以下で安定に保たれるが,pH6では完全に失活した。また活性は温度の上昇と共に減少し,50℃以上では30分間で完全に失活した。感受性酵母に対するSMKトキシンの見かけのキラー活性はアッセイ培地のNaCl及びKClの濃度の増加に伴って増大した。 SMKトキシン活性精製画分の酸性条件下でのポリアクリルアミドゲル電気泳動及びトリシンSDS-ポリアクリルアミドゲル電気泳動の結果から,SMKトキシンは,(4kDa)と(8kDa)のサブユニットからなる2量体であることが示された。両サブユニット間にはジスルフィド結合は無く,酸性(pH4以下)から中性へのpH変化で両サブユニットは容易に解離すること,解離と同時にサブユニットは凝集して沈澱しサブユニットは溶液中に残留することが示された。この結果から,サブユニットはサブユニットとの結合を介して可溶性の性質を保持しているものと推定された。 第III章では,SMKトキシンをコードする遺伝子SMK1の解析について論じている。サブユニットのアミノ酸配列からプライマーを作製し,ゲノムDNAのPCR分析を行った。得られたPCR断片をプローブとして,ゲノムDNAライブラリーから5.5-kbのEoo RI断片のクローン化に成功した。この断片に含まれる1.4-kbのHind III断片の塩基配列中には222アミノ酸をコードする666-bpのオープンリーディングフレーム(ORF)が存在した。及びサブユニットのN末端アミノ酸配列分析及びサブユニットのC末端ペプチドの配列分析の結果から,このORFにはシグナルペプチド,サブユニット,ORF中で唯一の推定上のN結合型糖鎖を持つ介在配列rペプチド,及びサブユニットを含む前駆体がコードされており,およびサブユニットはそれぞれ63残基および77アミノ酸残基からなるペプチドとして切り出されることが示された。SMKトキシンとSaccharomyces cerevisiaeのK1トキシンとの間に1次構造上の類似性は認められないが,その前駆体構造,疎水性度,プロセシングの様式等において共通点が見いだされた。なお染色体電気泳動及びハイプリダイゼーションにより,SMK1が染色体上に存在することも確かめられている。 第IV章では,抗サブユニット抗体に反応する糖タンパク質Gp26について論じている。P.farinosa KK1株を6%食塩を含む培地で培養した培養濾液中には,ウェスタソブロットにより抗サブユニット抗体と反応する約26kDaのタンパク質(Gp26)が存在することが明らかになった。Gp26の分泌には0.5M以上の食塩を含むYPD(1%酵母エキス,2%ポリペプトン,2%グルコース)培地が必要であり,食塩濃度1.5-2.5MでGp26の分泌が最大になった。精製後,酵素消化等により,Gp26が糖タンパク質であることを確認するとともに,酵素消化断片のアミノ酸配列分析の結果から,Gp26はSMKトキシンのプロトキシンであることを示唆した。さらにノーザンブロット解析の結果.SMK1のmRNAは食塩非存在下でも定常的に転写されていることが示され,食塩により誘導されるGp26の分泌は転写以降の翻訳あるいは分泌経路の過程で制御されていることが推定された。 以上,本論文は耐塩性酵母Pichia farinosaの新規キラー因子(SMKトキシン)について,その構造,および遺伝子について検討した結果を論じたもので学術上,応用上貢献するところが少なくない。よって,審査員一同は,本論文が博士(農学)の学位論文として価値有るものと認めた次第である。 |