生物はさまざまなかたちで環境に適応しているが、その一つに微生物との共生がある。事実、少なからぬ動植物は自身にはない微生物の代謝系を利用している。このような共生現象の中でも、微生物が宿主の細胞内にまで入り込んだ細胞内共生は宿主-微生物間の相互作用がきわめて強いものと考えられる。細胞内共生は、根粒菌を獲得したマメ科植物での窒素固定、海産無脊椎動物の藻類との共生による光合成など機能的多様性に富んでいる。陸生動物ではゴキブリ目、半翅目を中心として昆虫類の約10%が細胞内共生微生物を保有していることが知られている。本研究ではアブラムシ細胞内共生系について、その機能および宿主-共生バクテリア間の物質交換に関する一連の実験を行った。 アブラムシは半翅目・同翅亜目に属し、栄養的に偏りの大きい植物師管液を常食としながらも極めて旺盛な繁殖力を持つ。年に一度の両性生殖をのぞけば、アブラムシの生殖は二倍体性の単為生殖に基づき、この世代の虫は盛んに幼虫を産み続ける。アブラムシの共生微生物(共生体)は大腸菌に類縁度の高いバクテリアで、菌細胞という脂肪体の中の巨大な遊離細胞に収納されている。抗生物質処理などにより共生体を除去すると虫の成長は著しく阻害されると同時に、この虫の特徴である旺盛な繁殖力が失われる。植物師管液は糖分を豊富に含む反面、窒素に乏しい。師管液中の窒素はもっぱら遊離アミノ酸として存在しているが、いわゆる必須アミノ酸が少なく動物の餌としてはアンバランスな組成になっている。これらのことから、アブラムシは成長と生殖に必要な窒素を共生体に依存していると考えられる。 実験には、ソラマメ(Vicia faba)の芽生えで飼育しているエンドウヒゲナガアブラムシ(Acyrthosiphon pisum)を用い、非共生個体はリファンピシンを注射した親虫から得た。本研究に先立つ研究で、ソラマメ師管液とアブラムシの排出物である甘露のアミノ酸分析を行った。師管液にはアスパラギン、グルタミンが多く、これら2つのアミド基を持つアミノ酸(アミド)が全体の50%近くを占めていた。共生個体と非共生個体の甘露を比較したところ、後者ではアミド、特にグルタミンの含有量が高く、共生体の存在により師管液中に相対的に他のアミノ酸より多く含まれるアミドの利用度が上がることが示唆された。 本研究では、アブラムシ窒素代謝における共生体の役割をさらに詳しく調べる目的で、虫を人工飼料で飼育した。飼料はアミノ酸組成を師管液に似せたものと、虫の全組織のアミノ酸組成に基づき必須アミノ酸を充分に含むものを用意し、共生個体と非共生個体の成長および産仔を植物上と2つの人工飼料上で比較した。共生個体はすべての飼料で産仔能力があったが、非共生個体は植物とアミノ酸組成を師管液に似せた人工飼料では成虫にはなったものの完全に不妊で、アミノ酸バランスのよい人工飼料でのみ次世代を残した。このことは、アブラムシが偏ったアミノ酸組成をもつ植物師管液を常食としながら繁殖するために共生体が不可欠であることを意味している。さらに、人工飼料にアミド基が15Nで標織されたグルタミンを導入し、虫のアミノ酸合成能力を調べたところ、いわゆる必須アミノ酸は共生個体でのみ合成されていた。共生体の機能の一つは、師管液中の主要な窒素源であるアミドを利用して必須アミノ酸を合成し、アンバランスなアミノ酸組成を補正することであると考えられる。 窒素代謝における共生体のもう一つ別の機能として、虫の代謝老廃物の再利用が挙げられる。ところが、他の多くの昆虫とは異なりアブラムシは尿酸を合成せず、この虫の窒素代謝老廃物が何であるかは不明であった。非共生個体は多量のアミド、特にグルタミンを含んでいたが、これらのアミドは単に飼料の一部が利用されずに捨てられただけでなく、虫の代謝老廃物として排出されたものであるとも考えられた。そこでアミドを含まない人工飼料で虫を飼育し、甘露のアミノ酸分析を行った。飼料中のアミドの有無に関わらず、共生個体はアミドをほとんど排出せず、非共生個体は多量のアミドを排出した。特にグルタミンは、これを含まない飼料で飼育した場合でも、非共生個体の甘露に最も多く含まれるアミノ酸であった。グルタミン合成酵素活性は、共生、非共生個体に差はなかった。窒素代謝により生じるアンモニアはグルタミン酸に取り込まれグルタミンのアミド基になることで処理され、非共生個体では代謝老廃物として捨てられるが、共生個体では再利用されると考えられる。また、非共生個体をアミド基が15Nで標識されたグルタミンを含む人工飼料で飼育すると、その甘露に含まれるアスパラギンとニンヒドリン反応陽性の未知化合物に高い標識がみられた。この化合物はNMR、MSおよびHPLCにより、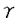 -グルタミルグルタミンと-グルタミルアスパラギンの混合物であると同定された。多くの動植物組織において、-グルタミルアミノ酸は-グルタミルトランスペプチダーゼにより合成されることが知られている。この細胞膜に結合した酵素は細胞内へのアミノ酸輸送に関与していることが示唆されており、アブラムシ細胞内共生系においてもアミノ酸代謝において重要な役割を果たしていると考えられる。 -グルタミルグルタミンと-グルタミルアスパラギンの混合物であると同定された。多くの動植物組織において、-グルタミルアミノ酸は-グルタミルトランスペプチダーゼにより合成されることが知られている。この細胞膜に結合した酵素は細胞内へのアミノ酸輸送に関与していることが示唆されており、アブラムシ細胞内共生系においてもアミノ酸代謝において重要な役割を果たしていると考えられる。 以上の実験からアブラムシ共生系ではアミドが窒素担体として機能し、共生体はそれを利用し虫自身では合成できない必須アミノ酸を生産することが示唆された。これは、グルタミンとアスパラギンが体液中の遊離アミノ酸の50%以上を占めること、外科的に虫から単離した菌細胞がグルタミンを盛んに取り込むこと、からも裏付けられた。ところが、菌細胞を破壊して得られる単離共生体はグルタミンをほとんど取り込まず、代わりにグルタミン酸をよく取り込んだ。菌細胞に取り込まれたグルタミンはその細胞質でグルタミン酸とアンモニアに加水分解された後に共生体に取り込まれると考えられる。実際、菌細胞に14C-グルタミンを取り込ませると、速やかにグルタミン酸に転換された。またアミド基が15Nでラベルされたグルタミンを取り込ませると、その細胞質に15N-グルタミン酸が形成され、グルタミンの加水分解により生じるアンモニアの一部は -ケトグルタル酸との反応によりグルタミン酸になることが示された。この反応はグルタミン酸デヒドロゲナーゼによると考えられる。これらを踏まえて、単離した共生体のアミノ酸合成を15N-グルタミン酸を用いて調べた。その結果、少なくとも4つの必須アミノ酸、すなわちイソロイシン、ロイシン、フェニルアラニンおよびバリンがグルタミン酸のアミノ基窒素を使って合成されることが明らかになった。 -ケトグルタル酸との反応によりグルタミン酸になることが示された。この反応はグルタミン酸デヒドロゲナーゼによると考えられる。これらを踏まえて、単離した共生体のアミノ酸合成を15N-グルタミン酸を用いて調べた。その結果、少なくとも4つの必須アミノ酸、すなわちイソロイシン、ロイシン、フェニルアラニンおよびバリンがグルタミン酸のアミノ基窒素を使って合成されることが明らかになった。 本研究では、アブラムシは共生体の持つアミノ酸合成能を利用した独特の代謝系を確立することにより師管液にうまく適応していることが示された。またアブラムシ共生系の宿主-共生体間の物質交換は、いくつかの点で、他の細胞内共生あるいは細胞内寄生に共通していた。アブラムシ共生体はグルタミン酸を盛んに取り込み他のアミノ酸合成の窒素供与体としていたが、グルタミン酸はリケッチアなどの細胞内寄生バクテリアの主要な呼吸基質であることが知られている。利用のされ方こそ異なれ、細胞内バクテリアの代謝においてグルタミン酸は重要な役割を担っているようである。単離されたアブラム共生体はグルタミンをほとんど取り込まなかった。このことは共生系の物質交換が選択的に行われていることを示唆している。マメ科植物の根粒においても根粒バクテロイドが利用できる化合物は限られ、バクテロイドを覆う宿主由来の膜がそれを制御している。アブラムシ共生体も宿主の膜に包まれており、この膜が選択的透過性を持っている可能性が高い。アブラムシ共生体をはじめ、細胞内に共生あるいは寄生するバクテリアは多量のストレス蛋白質を合成している。細胞内で利用できる物質が宿主の膜により制限されることと何らかの関係があるのかもしれない。さまざまな細胞内共生あるいは細胞内寄生はそれぞれの表現型が全く異なるが、そこには共通の生物学的機構が存在するように思われる。その機構は、個々の実例の詳細な解析を通じて明らかになると期待される。 |