本論文は4章からなり、第1章では色素体核(色素体のDNA-タンパク質複合体)の単離とそのキャラクタライゼーションについて、第2章では単離色素体核を用いたin vitro DNA合成系の開発と原色素体増殖過程における色素体ゲノムの複製制御について、第3章では単離色素体核を用いたin vitro転写系の開発と葉緑体分化過程における色素体遺伝子の転写制御について、第4章ではタバコ培養細胞におけるアミロプラスト分化誘導系の開発とアミロプラスト形成過程における色素体ゲノムの転写・複製機能の変化について述べられている。 生命の遺伝情報を担うDNAは、生体内ではタンパク質との相互作用により高次に組織化された状態で存在する。これらDNA-タンパク質複合体の構造変化は、ゲノムの複製や遺伝情報の発現に多大な影響を与えるものと考えられているが、確証は得られていない。色素体DNAもタンパク質と結合し、コンバクトに組織化された「色素体核」としてその機能を発現している。色素体核の構造と色素体ゲノムの転写・複製機能は、色素体分化過程で著しく変動することが示唆されている。また、色素体のゲノムサイズは120-180kbpと小さい。従って、色素体分化にともなう色素体核の構造と機能の変化は、DNA-タンパク質複合体の構造と転写・複製機能の関係を解析するうえで理想的なモデル系になり得る。論文提出者は、こうした観点から、色素体核の構造-機能連関の解析を目標として研究を行った。 色素体核の構造と機能の関連を解析するためには、構造と転写・複製機能を保持した色素体核の単離が必須であった。論文提出者は、第1章で、タバコ培養細胞BY-2の原色素体から、(1)高次構造の保持に重点をおく新しい方法と、(2)転写活性のみを指標とする従来の方法の2通りの単離法に従って、それぞれ「単離色素体核」および「Transeriptionally Active Chromosome(TAC)」と呼ばれるDNA-タンパク質複合体を単離し、その形態を比較した。その結果、単離色素体核は、DAPI蛍光顕微鏡観察によりコンパクトな蛍光スポットとして、また電子顕微鏡観察によりヌクレオソーム様のビーズ構造が三次元的に折り畳まれた構造として観察され、その高次構造が保持されているのに対し、TACは核構造が分散しており、高次構造が保たれていないことを明らかにした。さらに、単離色素体核の単位DNA当たりのUTP取り込み活性はTACの約5倍、dCTP取り込み活性は約50倍と極めて高く、単離色素体核はその高次構造だけでなく、in vivoにおける転写・複製機能もより良く保持している可能性が高いことを示した。 第2章では、単離色素体核のdCTP取り込み活性を詳細に解析し、色素体ゲノムのDNA合成能を測定するin vitroアッセイ系を開発した。単離色素体核は、膜画分の大部分と可溶性成分を失っているにも関わらず、単離色素体に匹敵する高いdCTP取り込み活性を示す。しかも、単離色素体の状態では内在性のDNase活性が検出されるのに対し単離色素体核にはそうした活性は検出されない。DNaseの混入がなく、高いdCTP取り込み活性を保持している単離色素体核は、DNA合成活性の測定に適していることが明らかになった。さらに、各種阻害剤処理や反応条件に対する応答、反応産物やDNAポリメラーゼの見かけの分子量の測定等により、単離色素体核のdCTP取り込みが、色素体に特異的な -like DNAポリメラーゼによる色素体DNAの合成反応であることを明らかにし、色素体ゲノムのDNA合成速度を測定するin vitroアッセイ系を確立した。論文提出者はさらに、この新しいアッセイ系を用いてタバコ培養細胞BY-2の増殖過程における色素体ゲノムの複製制御を解析している。定常期に入ったBY-2細胞を新しい培地に植え継ぐと、細胞は増殖を再開する。植え継ぎ後約12時間目から72時間目までが対数増殖期、96時間目以降が定常期に相当する。DAPI蛍光顕微鏡観察および定量的サザンハイプリダイゼーションの結果、植え継ぎ後24時間以内、細胞増殖のごく初期に色素体DNA量が著しく増加することが示された。単離色素体核のDNA合成活性は植え継ぎ後24時間以内に約5倍の活性化を示し、細胞増殖のごく初期に色素体ゲノムの複製が一過的に活性化されることが明らかになった。 -like DNAポリメラーゼによる色素体DNAの合成反応であることを明らかにし、色素体ゲノムのDNA合成速度を測定するin vitroアッセイ系を確立した。論文提出者はさらに、この新しいアッセイ系を用いてタバコ培養細胞BY-2の増殖過程における色素体ゲノムの複製制御を解析している。定常期に入ったBY-2細胞を新しい培地に植え継ぐと、細胞は増殖を再開する。植え継ぎ後約12時間目から72時間目までが対数増殖期、96時間目以降が定常期に相当する。DAPI蛍光顕微鏡観察および定量的サザンハイプリダイゼーションの結果、植え継ぎ後24時間以内、細胞増殖のごく初期に色素体DNA量が著しく増加することが示された。単離色素体核のDNA合成活性は植え継ぎ後24時間以内に約5倍の活性化を示し、細胞増殖のごく初期に色素体ゲノムの複製が一過的に活性化されることが明らかになった。 第3章では、単離色素体核のUTP取り込み活性が色素体特異的なRNAポリメラーゼによる転写反応であることを明らかにし、色素体ゲノムの転写活性を測定するin vitro転写系を確立した。さらに、タバコ成熟葉の葉肉細胞の葉緑体と、タバコ培養細胞BY-2の原色素体における色素体ゲノムの機能を比較解析した。葉肉細胞から単離した葉緑体核のDNA合成活性はBY-2から単離した原色素体核の1/3程度と低かったが、逆に、転写活性は約15倍の値を示した。葉緑体核と原色素体核では個々の色素体遺伝子の相対的な転写速度も変化していた。論文提出者は、こうした色素体遺伝子の転写活性の変化と転写産物蓄積量の変化の間に一定の相関がみられることを示し、葉緑体分化過程では個々の色素体遺伝子の転写制御が転写産物蓄積量の決定、ひいては色素体遺伝子の発現制御において重要な役割を果たしていることを示唆した。 第4章では、タバコ培養細胞BY-2において原色素体からアミロプラストへの分化を誘導する実験系を開発し、分化にともなう色素体ゲノムの機能変化を経時的に解析している。定常期のBY-2細胞を、オーキシン(2,4-D)を含まずサイトカイニン(BA)を添加した改変培地に植え継ぐと、細胞はほとんど分裂・増殖せず、色素体は大量のデンプンを蓄積して48時間以内にアミロプラスト化する。この過程では細胞当たりの色素体数、色素体DNAコピー数もほぼ一定であり、単離色素体核のDNA合成活性も顕著な上昇を示さない。単離色素体核の転写活性は植え継ぎ後急速に低下する。多くの色素体遺伝子の転写は植え継ぎ後24時間でほとんど停止してしまい、その転写産物は増加を停止する。例外的に、psbA(光化学系II反応中心32kDaタンパク質遺伝子)とpsaA/B(光化学系I反応中心P700アポタンパク質A1/A2遺伝子)の転写活性は培養過程を通じて比較的高く保たれており、その転写産物は増加し続ける。転写・複製機能の低下と共に、多くの色素体遺伝子の転写活性が低下するにも関わらず転写産物蓄積量はそれほど減少しないことから、アミロプラストでは転写産物の分解速度が極めて遅くなっていることが示唆された。 こうした一連の解析を通して、論文提出者は常に複数の手法を併用することにより、新たに開発した単離色素体核のin vitroアッセイ系がin vivoにおける色素体ゲノムの機能変化を再現し得ることを確認しつつ、原色素体、葉緑体、アミロプラストにおける色素体ゲノムの転写・複製機能の変化を明らかにし、さらに、サウスウェスタンプロッティング法により、色素体分化にともなって色素体核を構成するDNA結合タンパク質が変動することも示した。こうした一連の研究により、単離色素体核が高次構造、転写・複製機能を保持したDNA-タンパク質複合体であることを明らかにし、色素体ゲノムの機能制御機構を色素体核の構造変化という観点から解析するための基礎を築いた本論文提出者の業績は優れている。なお、本論文の第1、2、3、4章は河野重行、黒岩常祥両氏との共同研究であるが、論文提出者が主体となって解析および検証を行ったもので、論文提出者の寄与が充分であると判断する。従って、博士(理学)を授与できると認める。 | 
 m。 / 図2.単離色素体核のin vitro DNA合成産物とポリメラーゼの解析。 A:反応産物の解析。単離色素体核に[
m。 / 図2.単離色素体核のin vitro DNA合成産物とポリメラーゼの解析。 A:反応産物の解析。単離色素体核に[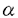 -32P]dCTP存在下でDNA合成反応を行わせた。反応後DNAを制限酵素HindIUで切断、アガロースゲル電気泳動後、(a)EB染色、(b)オートラジオグラフィーを行った。何れも色素体DNAに特徴的なバンドバターンを示す。 B:ポリメラーゼの解析。単離原色素体核タンパク質を変性サケ精子DNAを含むSDSポリアクリルアミドで電気泳動後renatureし、[
-32P]dCTP存在下でDNA合成反応を行わせた。反応後DNAを制限酵素HindIUで切断、アガロースゲル電気泳動後、(a)EB染色、(b)オートラジオグラフィーを行った。何れも色素体DNAに特徴的なバンドバターンを示す。 B:ポリメラーゼの解析。単離原色素体核タンパク質を変性サケ精子DNAを含むSDSポリアクリルアミドで電気泳動後renatureし、[