| 内容要旨 | | 緒言 生体の薬物に対する反応には大きな個体差が見られることがあり,その成因の1つに生体の薬物代謝能の変動を挙げることができる。薬物の代謝は多くの要因により影響を受けることが知られているが,最近では遺伝的な素因の重要性が判ってきた。ヒトにおいては,デブリソキンやメフェニトインの代謝反応に遺伝的に代謝能の低いpoor-metabolizer(PM)の存在が報告され,次いで多くの薬物の代謝にもPMの存在が指摘されるようになった。これらの遺伝的代謝多型が原因で副作用の発現や効果の減弱等様々な問題が生じている。薬物を開発する上で,患者に対してより効果的で安全性の高い治療を施す為に,代謝により生じる個体差の程度と成因,特に遺伝的多型との関係を調べることは非常に重要なことである。一方,医薬品開発には種々の実験動物が使用されているが,実験動物における薬物の体内動態の個体差を,代謝多型の点から詳細に報告した例はない。著者は抗アレルギー作用を示す(+)-4-[4-(4-methyl-phenyl)phenylmethoxy-1-piperidinyl]butyric acid((+)-MPPB,図1)がラットにおいて明らかな体内動態の個体差を示すことを見出し,個体差と遺伝的代謝多型との関連を明らかにすることを目的として研究を行った。  図1 MPPBの化学構造1.ラットにおけるクリアランスの差の検討1)(+)-MPPBのSD系ラットにおける体内動態 図1 MPPBの化学構造1.ラットにおけるクリアランスの差の検討1)(+)-MPPBのSD系ラットにおける体内動態 (+)-MPPBをSD系雄性ラットに経口あるいは静脈内投与した時,明らかな血漿中濃度の個体差が認められ,slow-metabolize(SM)群とrapid-metabolize(RM)群の2つの表現型(フェノタイプ)に分けることができた(図2)。この時,SMとRM群間に中間的な血漿中濃度及び尿中排泄率を示す個体は存在しなかった。両群の薬物動力学的パラメーターを比較すると消失半減期で約6倍,また,全身クリアランスで約10倍の差が認められた。 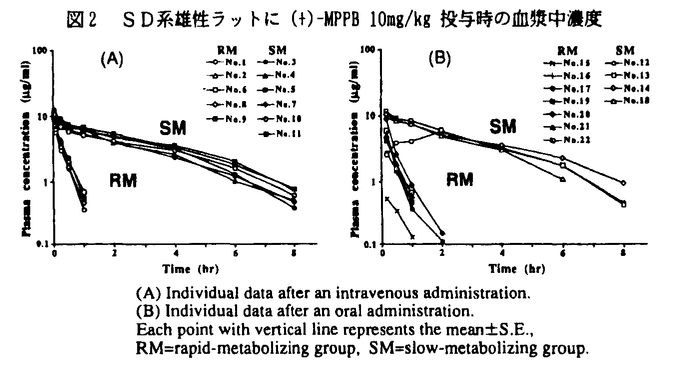 図2 SD系雄性ラットに(+)-MPPB 10mg/kg投与時の血漿中濃度 図2 SD系雄性ラットに(+)-MPPB 10mg/kg投与時の血漿中濃度 次に,個体差の原因が代謝能の差によるものと推察し,あらかじめフェノタイプを確認したSD系雄性ラットを用い,肝における(+)-MPPBの抽出率を定常状態法により求めた。得られた抽出率により算出された肝クリアランスはSM群で0.22±0.14liter/hr/kg,またRM群では2.47±0.08liter/hr/kgと,RM群が約10倍大きい値を示し,全身クリアランスの差は肝クリアランスの差に起因することが判った。 2)種々の系統における(+)-MPPB投与後のフェノタイプ 各種系統ラットに(+)-MPPBを経口投与し,得られた血漿中濃度よりそれぞれのフェノタイプを確認した。クローズドコロニーであるSD系雄性ラットは入手先にかかわらず,SM群とRM群が混在した.また,SD系雌性ラットも同様に混在型であり,雌雄差は認められなかった。SD系と同じくクローズドコロニーであるWistar系も混在型であった。一方,クローズドコロニーに比べ遺伝的により純粋である近交系のLewis系は全てがRM群,一方,Fischer344系及びACI系は全てがSM群であった。すなわちクローズドコロニーはいずれもRM群とSM群の混在型であり,一方,近交系では全個体が必ずどちらか一方のフェノタイプであることが確認され,明らかに遺伝的な素因が関与していると考えられた。 3)光学異性体の体内動態 光学異性体である(-)-MPPBの体内動態を明らかにする為に,SD系雄性ラットに(-)-MPPBを経口投与し血漿中濃度を調べたところ,(-)-MPPB投与時においても(+)体と同様の明らかな個体差が存在することが判り,さらに両光学異性体におけるフェノタイプは完全に一致していた。 2.代謝経路に関する検討1)肝ミクロソームによる(+)-及び(-)-MPPBのin vitro代謝 フェノタイプを確認したSD系雄性ラットから調製した肝ミクロソームを用いた検討において,(+)-MPPBを基質とした時には代謝物M1が,また(-)-MPPBを基質とした時には代謝物M1及びM2が検出され,両異性体間で代謝経路の違いが認められた(図3)。これらの代謝物はRM群に比べSM群では非常に生成量が少ないか又はほとんど検出されず,M1及びM2への代謝能の差が個体差の原因と考えられた。  図3 ラット肝ミクロソームによるMPPBの代謝経路 図3 ラット肝ミクロソームによるMPPBの代謝経路 次に,肝ミクロソームによる(+)-MPPBのM1への代謝速度を基質濃度を変え測定し,それぞれのKm,Vmaxを算出した。その結果,RM群では高親和性(Km=4.4〜6,6 M)及び低親和性(Km=471〜577M)の2種類の酵素が関与し,一方,SM群では低親和性(Km=304〜510M)の酵素のみが関与していた。両酵素の代謝固有クリアランスを比較すると,高親和性酵素が低親和性のものに比べ値が約100倍大きく,高親和性酵素が肝ミクロソームのMPPB代謝のほとんどに寄与していることが判った。また,阻害剤を用いた検討の結果,いずれの酵素もチトクロームP-450であることが判った。 M)及び低親和性(Km=471〜577M)の2種類の酵素が関与し,一方,SM群では低親和性(Km=304〜510M)の酵素のみが関与していた。両酵素の代謝固有クリアランスを比較すると,高親和性酵素が低親和性のものに比べ値が約100倍大きく,高親和性酵素が肝ミクロソームのMPPB代謝のほとんどに寄与していることが判った。また,阻害剤を用いた検討の結果,いずれの酵素もチトクロームP-450であることが判った。 以上より,MPPBのラット肝ミクロソームによる代謝には親和性の異なる2種類のチトクロームP-450が関与しており,さらに高親和性の分子種は多型性を有することが判った。SM群のラットには代謝能のほとんどを占める高親和性チトクロームP-450が欠損している為にMPPBのラットの体内動態において個体差が現れると考えられた。また,この多型性を有するチトクロームP-450はMPPB光学異性体を異なる経路で代謝することも明らかとなった。 2)In vivoにおける(+)-MPPBの代謝 フェノタイプを確認したSD系雄性ラットに(+)-MPPBを経口投与後,尿及び胆汁を採取し,Frit-FAB LC/MSを用い代謝物のプロファイルを検討した。その結果,RM群では胆汁及び尿中のいずれにも未変化体は検出されず,胆汁中においてin vitro試験では検出されなかった代謝物M3が検出された。M3はFAB-マススペクトルの結果から,M1がさらに酸化されたカルボキシル基を有する代謝物と推定された。一方,SM群では未変化体が胆汁,尿中の両方に認められた。さらに尿中においてbutyric acid側鎖が 酸化された代謝物が認められ,代償的な代謝反応が進行しているものと考えられた。以上のように,in vivoにおいても代謝による個体差を確認することができた(図4)。 酸化された代謝物が認められ,代償的な代謝反応が進行しているものと考えられた。以上のように,in vivoにおいても代謝による個体差を確認することができた(図4)。  図4 ラットにおける(+)-MPPBの代謝経路3.代謝多型と構造活性相関 図4 ラットにおける(+)-MPPBの代謝経路3.代謝多型と構造活性相関 代謝多型による個体差の程度と化学構造との関係を調べる為に,MPPBの構造類似体(ラセミ体)を合成し,SM及びRM群の肝ミクロソームによる代謝反応速度を比較検討した。 1)フェニル基上の置換基の効果 MPPBの4位メチル置換基を変換し,SM及びRM群の肝ミクロソームによる代謝速度を基質の残存率から比較した。その結果,置換基の位置,側鎖の炭素数の違い,またはハロゲン化等の変換によっても程度の差はあるものの,すべてにおいてSMとRM群間で代謝速度の差が認められた(P<0.01)。 2)双極性基側鎖の変換効果 MPPBのN-カルボキシル基間の炭素数を変え,同様に代謝反応速度を比較した。その結果,炭素数n=5まではMPPBと同様にSM群に比べてRM群で明らかに代謝速度が速く差が認められたが,n≧6では代謝速度が逆転した。これはN-カルボキシル基間の炭素数が長くなるにつれて,SM群のラットにおいてはpolymorphicなP-450が関与しない別の代謝反応の寄与率が大きくなる為と考えられた。 4.生理学的モデルによる体内動態の解明 これまでに得られたパラメーターを基に,非線形な代謝過程を組み込んだ生理学モデルを構築し,(+)-MPPBのSD系RM群ラットにおける体内動態の定量的な解析を試みた。 1)生理学的モデルによる血漿中濃度の予測 (+)-MPPBの生理学的モデルを構築し,(+)-MPPBを0.1-50mg/kgで経口及び静脈内投与した時の血漿中濃度の予測を行った。その結果,10mg/kgで投与後の実測値と予測値はほぼ一致し,in vitroで求められた代謝パラメーターによりin vivoの体内動態を予測できることが明らかとなった(図5)。  図5 血漿中濃度の予測2)初回通過効果における飽和 図5 血漿中濃度の予測2)初回通過効果における飽和 上記生理学的モデルに従い,投与量及び吸収速度(Ka)を変化させB.A.(F)を予測した。その結果,投与量を0.1-50mgで変化させた時,及びKaを変化させた場合のいずれもF値が大きく変化し,(+)-MPPBを10mg/kgで投与時に初回通過効果の飽和が生じていることを示すことができた。 結語 本研究では,ラットにおける薬物体内動態の個体差と遺伝的代謝多型との関連について検討を行った。 抗アレルギー作用を有する(+)-MPPB又はその光学異性体である(-)-MPPBがラットにおいて体内動態の個体差を示すことを見出した。この原因は,代謝物の同定及び酵素学的な検討からMPPBの代謝に関与するチトクロームP-450の代謝能の差に起因することを明らかにすることができ,遺伝的代謝多型の可能性が示唆された。実験動物において同一系統,同一性であるにもかかわらず,薬物の体内動態に遺伝的代謝多型に基づく個体差が存在することを初めて示すことができた。このことは実験動物としてのラットの使用に十分な注意を喚起するものである。この多型性を有するチトクロームP-450は光学異性体間で異なる代謝物を生成することを明らかにした。さらに,MPPBの種々の構造類自体を用いた代謝実験により,個体差が生ずる範囲について興味ある結果を得ることができた。今回,in vitro代謝試験で得られた代謝速度に関するパラメーターを用い,生理学的モデルによる体内動態の解析を行った。その結果,非線形kineticsを組み込んだ生理学的モデルによる解析の有用性が示され,特に初回通過効果の飽和現象を定量的に裏付けることができた。 |