エンドセリン-1(ET-1)は、1988年に培養血管内皮細胞の上清より得られた強力な血管収縮物質として報告された21アミノ酸残基よりなるペプチドで、ET-2,3とともにペプチドファミリーを形成している。ET-1は、平滑筋収縮作用のほか細胞増殖・ホルモン分泌の制御・神経系の調節など、広範囲な生物活性をもつことが報告されてきているが、その生理的役割や循環器疾患をはじめとする各種の病態における意義はまだ明らかにされていない。一方、1988年に米国ユタ大学のCapecchiらが初めて成功させて以来、ジーンターゲティングと呼ばれる手法によって染色体の特定遺伝子に人為的に突然変異を導入した遺伝子欠損マウスを作ることが可能になった。その表現型を解析することにより、欠損した遺伝子の生理機能が明らかになるばかりでなく、ヒト疾患のモデルとして遺伝子診断や治療の開発への発展も期待できる。そこで、ET-1の生理学的、病態生理学的意義を明らかにするために、ジーンターゲティングの手法を用いてET-1欠損マウスを樹立した。 遺伝子欠損マウスは、マウスの胚盤胞の内細胞塊から株化された胚幹(ES)細胞に変異遺伝子断片を導入して、相同組み換えによって特定の染色体遺伝子に突然変異を起こさせ、これをマウスの胚盤胞に打ち込んでキメラマウスを作成し、変異遺伝子を受け継いだ子孫を得ることにより樹立される。まず、BALB/cマウス遺伝子ライブラリーよりマウスET-1遺伝子をクローニングし、第2〜5エクソンを含む7.5kbの断片のうちET-1の21残基アミノ酸をコードする第2エクソンにネオマイシン耐性遺伝子(neor)を挿入し、コンストラクト3’端に単純ヘルペス由来チミジンキナーゼ遺伝子(HSV-tk)を連結したDNAコンストラクトを作成した。このコンストラクトをエレクトロポレーションにより白色マウス129Sv/Jより樹立されたES細胞A3.1に導入し、G418 300 g/ml、Gancyclovir0.5Mでpositive-negative selectionを行った。この際、G418、Gancyclovirの至適濃度を検討するとともに、相同組み換えES細胞クローニングのtime courseを工夫した。G418・gancyclovir存在下で生き残った281クローンをPCR・Southem解析したところ、35クローンに一方の対立遺伝子で相同組み換えが起こったことが示された。さらにこの35クローンの核型を検索し、マウス正2倍体の40本の染色体をもつ細胞の率の高いクローンを黒色マウスC57BL/6Jより得られた胎盤胞に打ち込み、キメラマウスを得た。このうち独立した2系統のES細胞由来のキメラマウスでgermline transmissionが確認され、その子孫よりET-1遺伝子欠損マウスを樹立した。 g/ml、Gancyclovir0.5Mでpositive-negative selectionを行った。この際、G418、Gancyclovirの至適濃度を検討するとともに、相同組み換えES細胞クローニングのtime courseを工夫した。G418・gancyclovir存在下で生き残った281クローンをPCR・Southem解析したところ、35クローンに一方の対立遺伝子で相同組み換えが起こったことが示された。さらにこの35クローンの核型を検索し、マウス正2倍体の40本の染色体をもつ細胞の率の高いクローンを黒色マウスC57BL/6Jより得られた胎盤胞に打ち込み、キメラマウスを得た。このうち独立した2系統のES細胞由来のキメラマウスでgermline transmissionが確認され、その子孫よりET-1遺伝子欠損マウスを樹立した。 キメラマウスの交配により得られたET-1変異へテロ接合体マウスは外見・発育とも正常であった。ヘテロ接合体同士の交配により得られたホモ接合体(ET-1完全欠損型マウス)は出生直後に呼吸不全にて死亡した。さらに、図のように、下顎形成不全・小耳介・舌低形成など頭頚部に明らかな形成異常を認めた。組織学的検索では、ホモ接合体は前頚部の腺組織・結合織の発達が乏しく、口蓋裂を認め、外耳および中耳構造のほとんどが欠損していた。アリザリンレッド・アルシアンブルーによる骨軟骨染色では、下顎骨・側頭骨・舌骨・甲状軟骨などに著しい異常を認め、胎生期の経時的骨軟骨染色により、鰓弓由来のMeckel・Reichert軟骨の発達不全からこれらの骨格異常をきたしていることが明らかになった。 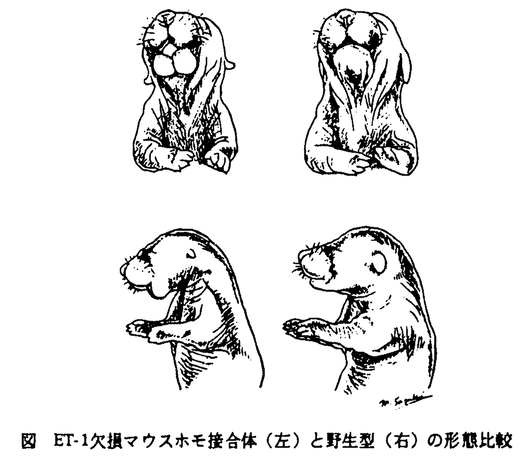 図 ET-1欠損マウスホモ接合体(左)と野生型(右)の形態比較 図 ET-1欠損マウスホモ接合体(左)と野生型(右)の形態比較 組織学的検索では、ホモ接合体は前頚部の腺組織・結合織の発達が乏しく、口蓋裂を認め、外耳および中耳構造のほとんどが欠損していた。アリザリンレッド・アルシアンブルーによる骨軟骨染色では、下顎骨・側頭骨・舌骨・甲状軟骨などに著しい異常を認め、胎生期の経時的骨軟骨染色により、鰓弓由来のMeckel・Reichert軟骨の発達不全からこれらの骨格異常をきたしていることが明らかになった。 ET-1はこれまで主として循環調節因子として数多くの研究がなされてきたものの、形態形成におけるET-1の関与については全く注目されていなかった。これを証明するため、胎生期におけるET-1の作用と発現部位を検討した。胎生9.5日の下顎弓の器官培養において、ホモ接合体で舌上皮の低形成がみられたが、ET-1はこの系で舌上皮発達を亢進させる作用を示した。また、胎生9.5日のwhole mount in situ hybridazationにおいて、ET-1mRNAの発現は鰓弓の上皮細胞に最も著明に認められた。これらの結果から、胎生期の鰓弓由来の組織・器官形成において、ET-1が重要な役割を担っていることが明らかになった。 一方、ET-1の循環調節における生理的あるいは病態生理的意義は最も興味深いところでであるが、ホモ接合体は生後直後に死亡するのでその循環動態の解析は不可能であった。そこで、ET-1の産生が約半分に低下しているヘテロ接合体と野生型マウスの循環動態を比較検討した。オスのマウスの大腿動脈に外径0.5mmのポリエチレンチューブを挿入し、観血的に無麻酔無拘束下またはウレタン麻酔下で動脈圧をモニターしてその動態を解析した。ET-1は強力な血管収縮物質であり、静脈内に投与すると長時間持続する血圧上昇をきたすことが知られており、また、ヘテロ接合体のET-1血中濃度は野生型の約半分であるので、ヘテロ接合体で血圧は低下すると思われたが、予想に反してヘテロ接合体の血圧はいずれの条件下でも野生型に比べて約10mmHgほどの高値を示した(ヘテロ接合体の収縮期、平均、拡張期血圧:133±2,116±2,99±2mmHg、野生型では、122±2,105±2,88±3mmHg)。心拍数には有意な差はなかった。 この結果について、ET-1の産生低下が受容体のup regulationなどによりET-1に対する反応性を亢進させている可能性を考え、ET-11nmol/kg i.v.に対する昇圧反応にはへテロ接合体と野生型で差がなかった(それぞれ32±3,32±3mmHg上昇)。また、ET-1が内皮細胞におけるNOの産生を促進することから、ヘテロ接合体における血圧上昇がNOの産生低下による可能性も考えられたが、NO合成酵素の阻害薬L-NMMA 250mol/kg投与後も血圧の差は変わらず(それぞれ35±3,35±4mmHg上昇)、これも否定的であった。 ジーンターゲティングによって樹立したET-1遺伝子欠損マウスがこのような予想外の表現型を示したことは、血管収縮・昇圧ペプチドとしてとらえられてきたET-1が、胎生期の形態形成において重要な生理的意義をもつ事を示しており、さらに、ET-1の循環調節における役割は違った面からも考える必要性を示している。 ホモ接合体の致死的呼吸不全の原因の一つは、頚部のドーナツ型の異常骨による気道の狭窄であるが、このほかにも、呼吸循環中枢におけるET-1の関与が重要であると思われる。また、ホモ接合体の頭頚部の奇形発生部位は、全て鰓弓由来組織であり、神経堤細胞由来間葉織の分化にET-1が関与していると考えられる。そして、これらの形態異常が、ET-1遺伝子以外の突然変異によって起こったとは考えにくい。なぜなら、遺伝子型と表現型が239匹の胎児または新生児において完全に一致し、独立した2種類の相同組み換えESクローンから同様のホモ接合体の表現型が得られていること、さらに、下顎培養においてホモ接合体の舌上皮の形成がET-1によって正常化し、胎児の鰓弓に一致してET-1の発現が見られることからも確かである。胚葉誘導や器官形成など発生現象の分子機構の研究は、TGF ファミリーなどの成長因子やホメオティック遺伝子などを中心に進められているが、まだ多くは解明されておらず、本研究もその端緒の一つとなるであろう。また、ホモ接合体の表現型はpierre-Robin症候群・Treacher-Collins症候群などの疾患と極めて似ており、これらの疾患の原因遺伝子解明や新しい治療法の開発への手掛かりとなる可能性が考えられる。 ファミリーなどの成長因子やホメオティック遺伝子などを中心に進められているが、まだ多くは解明されておらず、本研究もその端緒の一つとなるであろう。また、ホモ接合体の表現型はpierre-Robin症候群・Treacher-Collins症候群などの疾患と極めて似ており、これらの疾患の原因遺伝子解明や新しい治療法の開発への手掛かりとなる可能性が考えられる。 ヘテロ接合体における血圧上昇は、これまでの報告、即ちET-1の昇圧作用や血中ET-1と高血圧の相関などからは全く予想外の結果であった。その機序を考える上で、ET-1が循環ホルモンよりもむしろパラクリンあるいはオートクリン因子として循環調節に関与していると考えられること、ET-1の内皮細胞に対するNO産生促進作用や、中枢性の部位特異的な血圧降下作用などをも考慮する必要がある。しかし、L-NMMAの昇圧作用にはへテロ接合体と野生型で差がなく、NO産生の低下による可能性は否定的である。ヘテロ接合体の血圧上昇の機序についてはET-1の神経性呼吸循環調節における関与を含めた詳細な解析が必要である。今後その解析を進めていくことにより、循環器疾患の病態生理やET拮抗薬の臨床応用を考える上で多くの示唆を与えていくと思われる。 |