C型肝炎ウイルス(以下HCV)の塩基配列が同定されて以来、HCVに関する研究が多数発表されていろ。HCVは非A非B型肝炎の主たる原因ウイルスであり、慢性肝炎、肝硬変、肝臓癌の進展に関与している。HCVの構造と機能が主に分子生物学的手法を用いて解析されつつある。叉HCV排除目的にインターフェロンが臨床の場で用いられている。HCV感染に伴い肝細胞障害が惹起されるがその機序は明らかでない。一般にウイルス感染に伴い感染細胞が障害を受ける機序として、ウイルス自体のcytopathicな効果と宿主の免疫を介したウイルス感染細胞の排除が考えられる。A型肝炎、B型肝炎ウイルス感染において肝細胞障害の主体は細胞性免疫であると考えられている。HCV感染における細胞性免疫応答に関する報告が集積されつつある。このような状況において、我々はC型肝炎の病因における細胞性免疫の関与を検討するため、C型肝炎核蛋白(以下NP)特異的細胞障害性T細胞(以下CTL)を慢性C型肝炎末梢血リンパ球(以下PBL)より合成ペプチドを用いて誘導することを試みた。 慢性C型肝炎患者のPBLを4種類のHCV NP混合合成ペプチドで2回反復刺激することにより、いずれかの混合合成ペプチド添加標的細胞を特異的に融解するCTLが慢性C型肝炎患者10例中7例のPBLより誘導され、各症例において細胞障害活性の得られるペプチドグループに差異が認められた。以上より慢性C型肝炎患者末梢血におけるHCV NP特異的CTLの存在が示唆され、又同CTLの標的エピトープが複数存在する事が示唆された。 最も特異性の高い細胞障害活性を示した症例のPBLにペプチド刺激を一週間ごとに反復し、特異的細胞障害活性の推移を検討したところ、ペプチド刺激を一週間毎に2回加えた場合最も特異性の高い細胞障害活性を得る事ができると考えられた。CTLの認識する抗原エピトープがNP81-100残基(以下NP9)内にあり、NP9単独でも特異的CTLの誘導が可能であった。NP9誘導CTLはNP9添加標的細胞のみならずHCV NPを内因性に合成する組み換えVaccinia Virus感染標的細胞をも特異的に融解した。CTLのphenotypeがCD8陽性であることが示され、NP9の認識はHLA class I分子B44に拘束性があることが示された。又CTLの認識がT細胞受容体を介している事が示唆された。HCV NP9内に含まれるCTLエピトープのHLA B44 haplotypeにおける免疫学的優位性を評価するために、HLA B44分子を有する複数症例及び対照例を解析した。HLA B44分子を有する慢性C型肝炎患者6例中3例のPBLをNP9で刺激することによりNP9特異的CTLの誘導が可能であったが、一方HLA B44分子を有する健常例4例においてNP9で刺激されたPBLはいずれもNP9添加されたHLA B44分子を有する標的細胞に対する特異的細胞障害活性を示さず、HLA B44分子を有する慢性C型肝炎症例においてHCV感染とHCV NP特異的CTLの誘導性に密接な関連のある事が示唆された。 HLA B44拘束性HCV NP特異的CTLのエピトープを有するNP9の91アミノ酸残基を変化させた合成ペプチドを用いてCTLの誘導及び誘導されたCTLの解析を試みた。NP9の91アミノ酸をLeucineからMethionineに変化させてもCTLの誘導及び誘導されたCTLに対すろ標的抗原性に変化をきたさないが、一方91アミノ酸をCysteineに変化させたペプチドはCTLの誘導ができない上に誘導されたCTLに対する標的抗原性が消失することが判明した。ウイルス亜分類に対する細胞性免疫の多様性がウイルス感染においていかなる役割を果たしているのか検討するため、患者血清中に存在するHCVにおける同部位のアミノ酸配列をRT-PCR及びdirect sequenceにより解析した。HLA B44拘束性CTLの認識を受けるアミノ酸配列に相当するHCV亜種が血中に存在する症例の中に、特異的CTLの誘導される例と誘導されない例が存在した。 HLA B44拘束性NP9誘導性CTLの最小認識エピトープを解析する為、誘導されたCTLに対し、NP9のアミノ酸を短くしたペプチド特異的細胞障害活性測定したところ、抗原エピトープを有する最小アミノ酸配列はHCV NPアミノ酸残基88-96の9アミノ酸であることが示された。エピトープの抗原性に重要な91アミノ酸が抗原提示細胞のHLA B44分子に結合するのかCTLのT細胞受容体と結合するのかを検討する方法を考えた。阻害実験により91アミノ酸がHLA B44分子との結合に関与している可能性が示唆されたが、偽陰性の可能性もあり結果の判断は慎重を要すると考えられた。 HLA class I分子B44を有する3例の慢性C型肝炎患者PBLよりHCV NP特異的CTLを誘導する際、HCV NP5’側の120アミノ酸よりなる精製蛋白であるJCC-2抗原を加える事により誘導された細胞障害活性の増加する症例が示された。Interleukin-2源であるLymphocultT(以下LY-T)を無添加の場合JCC-2抗原の有無にかかわらず細胞障害活性は誘導されなかった。LY-Tを添加した場合誘導される細胞障害活性は濃度依存性に増強したが、各々のLY-T濃度においてJCC-2抗原添加に伴い誘導される細胞障害活性の増加が認められた。その細胞障害活性の増加がHCV特異的ヘルパーT細胞(以下Th)の誘導によるものであるかどうか検討するため、慢性C型肝炎患者PBLよりHCV特異的Thの誘導を試みた。分離されたPBLに対しLY-T非添加の条件下に外因性にHCV抗原蛋白を加える事により特異的細胞増殖の有無を検討した。この症例において、PBL中にJCC-2抗原特異的に増殖する細胞の存在が示唆された。次にPBLをin vitroでJCC-2抗原で一回刺激する事によりJCC-2抗原特異的細胞増殖を誘導し、誘導された細胞を解析したところ、JCC-2抗原誘導性細胞は、JCC-2抗原濃度依存性で非常に特異性の高い細胞増殖を示した。又fresh PBLと同様にJCC-2抗原のみならずNP21-40残基及びNP81-110残基に対応する合成ペプチド対し特異的細胞増殖が認められた。JCC-2抗原誘導性細胞は大部分がCD3陽性、CD4陽性の表面抗原を有していた。又、その細胞はJCC-2抗原添加により特異的に増殖したが、その培養上清中に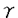 IFN活性が認められた。JCC-2抗原誘導性細胞は、NP9添加標的細胞に対する特異的細胞障害活性を認めずJCC-2抗原によりHCV NP特異的CTLが直接誘導される可能性は否定的である。上記の結果より、この症例のPBL中にHCV NP特異的CTLと共にHCV NP特異的Thが存在し、抗原特異的刺激によりHCV NP特異的CTLが誘導されろ際にJCC-2抗原特異的刺激を加える事により同時にHCV NP特異的Thが誘導され、HCV NP特異的Thの影響により誘導されたHCV NP特異的細胞障害活性が増強することがin vitroの実験系において示された。 IFN活性が認められた。JCC-2抗原誘導性細胞は、NP9添加標的細胞に対する特異的細胞障害活性を認めずJCC-2抗原によりHCV NP特異的CTLが直接誘導される可能性は否定的である。上記の結果より、この症例のPBL中にHCV NP特異的CTLと共にHCV NP特異的Thが存在し、抗原特異的刺激によりHCV NP特異的CTLが誘導されろ際にJCC-2抗原特異的刺激を加える事により同時にHCV NP特異的Thが誘導され、HCV NP特異的Thの影響により誘導されたHCV NP特異的細胞障害活性が増強することがin vitroの実験系において示された。 本研究において、下記の結果が得られた。 (1)慢性C型肝炎患者末梢血中におけるHCV NP特異的CTLの存在が示唆された。 (2)CD8陽性HCV NP特異的CTLがHLA class I分子B44を有する慢性C型肝炎患者より誘導され、その抗原エピトープが同定された。 (3)HLA B44拘束性CTLの認識する抗原エピトープ内のウイルス亜分類特異的な91アミノ酸残基が、CTLの抗原認識に重要な部位であった。 (4)HLA B44拘束性CTLが認識すろHCV NP領域内の最小認識エピトープが同定された。 (5)慢性C型肝炎患者末梢血中にHCV NP特異的Thが存在し、CTLと相互作用を示した。 C型肝炎における細胞性免疫に関する検討が、C型肝炎の病因解明に貢献すると共に特異的細胞性免疫の調節による治療へと結びつくことが期待される。 |