活性型ビタミンD(1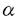 ,25(OH)2D3)は、主要なCa代謝調節ホルモンの一つであるが、同時に、破骨細胞の形成ばかりでなく、monocyte/macrophage系細胞の分化・増殖に重要な役割を果たしていることが知られている。一方、Macrophage-colony stimulating factor(M-CSF)はmonocyte/macrophageへの分化誘導および増殖を引き起こす因子として同定された糖蛋白であるが、破骨細胞の形成に必須の因子であることが示されている。さらに、1,25(OH)2D3とM-CSFはともに活性化されたマクロファージにより産生されることが知られている。しかしながら,1,25(OH)2D3とM-CSFの相互作用については未だほとんど研究されていなかった。 ,25(OH)2D3)は、主要なCa代謝調節ホルモンの一つであるが、同時に、破骨細胞の形成ばかりでなく、monocyte/macrophage系細胞の分化・増殖に重要な役割を果たしていることが知られている。一方、Macrophage-colony stimulating factor(M-CSF)はmonocyte/macrophageへの分化誘導および増殖を引き起こす因子として同定された糖蛋白であるが、破骨細胞の形成に必須の因子であることが示されている。さらに、1,25(OH)2D3とM-CSFはともに活性化されたマクロファージにより産生されることが知られている。しかしながら,1,25(OH)2D3とM-CSFの相互作用については未だほとんど研究されていなかった。 ヒトの単球やマクロファージが、M-CSFや1,25(OH)2D3などの刺激の下でin vitroにおいて、あるいはサルコイドーシスや慢性炎症の局所などにおいて、増殖することが明らかにされている。しかし、単球系細胞の増殖の機序については十分に明らかにされておらず、とくに、1,25(OH)2D3による単球の増殖の機序については研究されていなかった。申請者は、1,25(OH)2D3による単球増殖におけるM-CSFの役割について研究し、下記の結果を得ている。 (1)1,25(OH)2D3(10-8M)はヒト末梢血単球の6日間培養後の核の数を2.5倍に増加させたが、この増加は抗hM-CSF抗体(3.3mg/ml)によりほぼ完全に抑制された。しかし、抗hGM-CSF抗体(10mg/ml)、抗hIL-1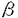 抗体(10mg/ml)、rabbit IgG(3.3mg/ml)は1,25(OH)2D3によるヒト末梢血単球の核の数の増加に影響を与えなかった。なお、抗hM-CSF抗体(3.3mg/ml)は対照群においては核の数に影響しなかった。 抗体(10mg/ml)、rabbit IgG(3.3mg/ml)は1,25(OH)2D3によるヒト末梢血単球の核の数の増加に影響を与えなかった。なお、抗hM-CSF抗体(3.3mg/ml)は対照群においては核の数に影響しなかった。 (2)抗hM-CSF抗体(3.3mg/ml)は1,25(OH)2D3(10-8M)やM-CSF(10ng/ml)による単球の増殖ほぼ完全に抑制したのに対し、rhGM-CSF(10ng/ml)やrhIL-3(10ng/ml)によるヒト末梢血単球の核数の増加は抑制しなかった。 (3)抗c-fms/M-CSF受容体抗体は、rhM-CSF(10ng/ml)によるヒト末梢血単球の核の数の増加を0.3mg/mlから3.0mg/mlまで用量依存的に抑制し、同様に、1,25(OH)2D3(10-8M)によるヒト末梢血単球の核の数の増加についても0.3mg/mlから3.0mg/mlまで用量依存的に抑制した。 (4)1,25(OH)2D3(10-8M)はヒト末梢血単球において48時間後の培養上清中のM-CSF濃度を対照に比べ約3倍に増加させた。 (5)TPA処理したTHP-1細胞、TPA処理したHL-60細胞において、1,25(OH)2D3(10-8M)は、蛋白レベル、mRNAレベルのいづれにおいてもM-CSFを、それぞれ、約3倍、約2倍に増加させた。、TPA処理したTHP-1細胞での1,25(OH)2D3によるM-CSF分泌促進については、用量依存的であり、かつ、種々のビタミンD誘導体のM-CSF分泌促進効果は、1,25(OH)2D3>>1(OH)D3>25(OH)D3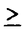 24R,25(OH)2D3の順であった。一方、CholecalciferolはM-CSF分泌に影響を認めなかった。 24R,25(OH)2D3の順であった。一方、CholecalciferolはM-CSF分泌に影響を認めなかった。 (6)最後に申請者は、1,25(OH)2D3による単球の増殖に単球自身が産生しているM-CSFが関与しているのかを確かめるために、M-CSFのantisenseを用いて核の数に対する影響を検討した。M-CSFのantisense(3.3mg/ml)は1,25(OH)2D3による単球の増殖を有意に抑えたが、sense、nonsenseは影響を及ぼさなかった。 以上、本論文は、1,25(OH)2D3によるヒト末梢血単球の増殖にはM-CSFが必要であること、そして、1,25(OH)2D3は、単球系細胞によるM-CSF産生を促進することを明らかにした。1,25(OH)2D3による単球増殖はM-CSF依存的であり、1,25(OH)2D3はM-CSF産生亢進を介して単球の増殖を惹き起している可能性が示された。 本研究は、1,25(OH)2D3濃度が高まっていることが知られているサルコイドーシス、結核、慢性炎症の局所などでのマクロファージの増殖の機序の解明に重要な貢献をなすものである。また、同時に1,25(OH)2D3による単球の増殖の機序、ならびに、活性型ビタミンDとM-CSFの間の相互作用ならびにcross talkの存在の可能性について新たな問題提起を投げかけたものといえる。 よって、学位の授与に値するものと考えられる。 |