2.B細胞分化におけるBCRの構造と機能 A.未熟Bリンパ球はIg-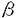 よりもIg-
よりもIg-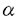 を過剩に発現している可能性がある:前述のCB3-1,CB3-2を用いることにより、Ig-の発現はpro-Bの段階から細胞質で始まり
を過剩に発現している可能性がある:前述のCB3-1,CB3-2を用いることにより、Ig-の発現はpro-Bの段階から細胞質で始まり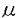 HCの膜発現と同時に細胞膜上に現われること、HCとIg-の膜発現量は厳密な相関関係にあることが明らかとなった。これに対し、Ig-のタンパクレベルでの発現はIg-のそれより早期に起こり、IgMとIg-の膜発現量の間には相関関係が認められないことが報告されている。sIgMの量はpre-BからBリンパ球に分化する過程で増加していくわけであるが、この結果はその際にsIgMとIg-の分子数比は常に一定であるのに対し、sIgMとIg-の比にばらつきがあることを意味する。 従って、BCRすなわちsIgM・Ig-・Ig-複合体の構成(分子数比)を考えた場合、sIgM:Ig-比はどのBCRでも一定であるのに対し、Ig-に関しては相対的にIg-の過剰なBCR、過小なBCRが存在し、未熟Bリンパ球のBCRにおいてはIg-がIg-に比し相対的に過剰に発現している可能性が示唆された。
HCの膜発現と同時に細胞膜上に現われること、HCとIg-の膜発現量は厳密な相関関係にあることが明らかとなった。これに対し、Ig-のタンパクレベルでの発現はIg-のそれより早期に起こり、IgMとIg-の膜発現量の間には相関関係が認められないことが報告されている。sIgMの量はpre-BからBリンパ球に分化する過程で増加していくわけであるが、この結果はその際にsIgMとIg-の分子数比は常に一定であるのに対し、sIgMとIg-の比にばらつきがあることを意味する。 従って、BCRすなわちsIgM・Ig-・Ig-複合体の構成(分子数比)を考えた場合、sIgM:Ig-比はどのBCRでも一定であるのに対し、Ig-に関しては相対的にIg-の過剰なBCR、過小なBCRが存在し、未熟Bリンパ球のBCRにおいてはIg-がIg-に比し相対的に過剰に発現している可能性が示唆された。
B.免疫グロブリン付随分子を介したBリンパ球の活性化は未熟Bリンパ球においてIg-優位で、成熟Bリンパ球においてIg-優位である:本論文において、Ig-を介したBリンパ球の活性化は成熟したBリンパ球においてのみ認められることを示した。抗Ig-抗体は主に未熟Bリンパ球を活性化し成熟Bリンパ球に対しては無効であることが報告されているが、抗Ig-抗体は逆に未熟Bリンパ球に対し無効で、成熟Bリンパ球に対しより効果的であった。この事実は、Ig-、Ig-を介したBリンパ球の活性化がその分化段階により異なることを示唆するものであり、上記Aで述べたIg-、Ig-の量的な差異を反映している可能性がある。
C.Ig-、Ig-の分子量・分子数はBリンパ球の分化段階、sIgのアイソタイプにより多様性がある:pre-Bリンパ球と成熟Bリンパ球のHCに付随するIg-とIg-の分子量には明瞭な差異が認められ、この分子量の違いは糖鎖修飾の違いによるものであった。一方、成熟Bリンパ球の各アイソタイプに付随するIg-のバンドはHCでは明瞭に、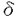 HCではやや不明瞭に認められたが、
HCではやや不明瞭に認められたが、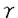 、HCに付随するIg-はそのバンドを同定出来なかった。その理由としては、 HCはHCと同様の分子量の大きな(糖鎖の多い)Ig-を付随しており、それが、HCの分子量とほぼ同じで単独のバンドとして検出できなかった可能性が高いと思われた。一方、Ig-に関しては、、、1、2、3HC全てでその存在が明らかで、その分子量には本文表4に示すような多様性が認められた。また、Ig-のバンドの濃さは、1、2、3HCにおいてHCに較べ明らかに薄かった。その理由についてはいくつかの可能性が考えられたが、フローサイトメーター上、HC陽性末梢血Bリンパ球のIg-の発現量が、HC陽性Bリンパ球よりも少なかったことを考慮すると(本論文図9)、、HC以外のアイソタイプには少数のIg-しか付随しないことが推察された。
、HCに付随するIg-はそのバンドを同定出来なかった。その理由としては、 HCはHCと同様の分子量の大きな(糖鎖の多い)Ig-を付随しており、それが、HCの分子量とほぼ同じで単独のバンドとして検出できなかった可能性が高いと思われた。一方、Ig-に関しては、、、1、2、3HC全てでその存在が明らかで、その分子量には本文表4に示すような多様性が認められた。また、Ig-のバンドの濃さは、1、2、3HCにおいてHCに較べ明らかに薄かった。その理由についてはいくつかの可能性が考えられたが、フローサイトメーター上、HC陽性末梢血Bリンパ球のIg-の発現量が、HC陽性Bリンパ球よりも少なかったことを考慮すると(本論文図9)、、HC以外のアイソタイプには少数のIg-しか付随しないことが推察された。
D.Ig-・Ig-ヘテロダイマーとsIgの結合親和性はsIgのアイソタイプにより異なる:本論文で、Ig-・Ig-ヘテロダイマーとIgMの結合はNP-40の存在下においても部分的には保たれるのに対し、IgDのそれは完全に失われることを示した。このことは、IgMとIg-・Ig-ヘテロダイマーとの結合親和性がIgDのそれよりも強いことが示唆する。
抗原のsIgへの結合はsIgの構造変化としてIg-・Ig-へ伝達されるが、その際sIgとIg-・Ig-の結合親和性の違いはシグナル伝達の効率の違いとして反映されるものと考えられる。また、sIg1分子あたりのIg-・Ig-の分子数に多様性があれば、仮に同一の強さのシグナルがsIgから伝えられても、ある場合はIg-優位のある場合はIg-優位のシグナルが細胞質内に入ることになる。Ig-、Ig-各々に接続する細胞内シグナル伝達系は異なっていることが知られており、sIgからの同一のシグナルが結果的に細胞質内へは量的・質的に異なったシグナルとして伝達されることが想像される。
本論文で、B細胞の分化段階の違いおよびHCアイソタイプの違いにおけるBCRの多様性を示し得た。今後の課題として、Ig-、Ig-の多様性特にHC:Ig-:Ig-の分子数比をより定量的に明らかにすると共にその多様性がIg-、Ig-に接続する細胞内シグナル伝達系に質的、量的にどのような影響を与えるのかを具体的に検討していく必要があるものと思われる。
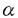 ,Ig-
,Ig-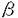 )の解析
)の解析