虚血心における側副血行路形成は急性虚血時の虚血性障害の軽減に重要な役割を果たすが、その形成機構については未だ不明な点が多く、血管新生因子の側副血行路形成への関与の可能性が注目されている。1987年,Leungらにより同定されたVascular endothelial growth factor(VEGF)は分泌性の血管内皮細胞特異的増殖因子で、その後多くの培養細胞、悪性腫瘍組織および心臓を含んだ正常組織において発現が認められることが報告されている。VEGFのタンパクとしての活性は様々なin vivo,in vitro実験により解明されつつあり、特に悪性腫瘍のin vivoでの増大を抗VEGE抗体が抑制すること、および一方で、虚血組織(ウサギ下肢阻血モデル)へのVEGF投与は側副血行路形成促進をもたらす等臨床応用的側面も示唆されている。しかしVEGFの発現調節については、細胞内メッセンジャーとしてcAMPおよびC-キナーゼの関与が言われているが、細胞外刺激については、卵巣において性周期の影響を受けること、悪性腫瘍で低酸素状態が発現を促すことが示唆されているのみで発現誘導の詳細なメカニズムは不明である。一方、卵巣以外の正常組織においては、その生理的あるいは病理的制御の有無については未解明である。そこで著者はVEGFの基礎的発現の認められる心臓において、虚血あるいは低酸素状態によりVEGFの発現が誘導されるかどうか、また発現誘導があった場合にその発現誘導機構を検討することにより、虚血心における側副血行路形成にかかわる可能性のある内因性因子の同定を試みることを研究目的とした。 (方法)ラットin vivoにおける冠動脈結さつモデルは3009程度のオスSDラットを麻酔人工呼吸下に開胸し直視下に左冠動脈を根部で結さつし閉胸、人工呼吸下においた。シャム手術として冠動脈結さつのみを除外した処置を行った。心臓灌流はオスSDラット(250g程度)から麻酔下に心臓を摘出し、素早くランゲンドルフ灌流装置に大動脈よりカヌーレーションし灌流を行った。虚血刺激としては一定時間の灌流停止を用いた。低酸素刺激としては灌流液のガス交換を通常の95%O2-5%CO2ガスの替わりに95%N2-5%CO2ガスを用い灌流液のPO2=40mmHgを得た。単離培養心筋細胞は3日令の新生ラット心室筋よりトリプシンによる分離を経て単離を行いコラーゲン塗付デイッシュで培養した。低酸素刺激はデイッシュを静置した小chamber中に95%N2-5%CO2ガスを吹き込むことでメデイウムPO2=40mmHgを得た。U-937細胞は通常の方法で継代し、低酸素刺激にはスターラー付のセミシールの培養ボトルを用いた。吹き込むガスのO2%をミキサーで調節することでPO2=40mmHgおよびPO2=90mmHgの二種類の低酸素状態を作成した。 RNAの調整はラット心組織よりはguanidinium thiocyanate methodで、培養細胞よりはacid guanidinium methodで行った。ラット、ヒトVEGF、ラットbasic fibroblast growth factor(bFGF)のcDNAはPCR法を用いて、それぞれの種由来のpolyadenylated RNAをreverse transcriptase処理して得たcDNAの第一鎖よりそれぞれに対応するプライマーの組み合わせて作成し塩基配列の確認を行った。ノーザンプロットは各実験ごとに等量の総RNAあるいはpolyadenylated RNAを変成、電気泳動、メンブレンへの転写後各cDNAより作成したラヂオラベルプローブでハイプリダイゼーションを施行、Fuji BAS 2000システムで分析した。 (結果)Study I.ラットin vivo冠動脈結さつモデルにおいて、冠動脈結さつラットの虚血側の左心室にのみ虚血後1時間でVEGF mRNAの強い誘導をみた。VEGF mRNAの発現は1-2時間をピークとして6時間までにコントロールレベルまで減少した。bFGFのシグナルは弱く、6時間までの経過中明確な発現は認められなかった。またランゲンドルフ灌流における虚血では10分前後の可逆的な虚血で虚血開始後30分で明確な誘導を示すことが示された。単離培養心筋における低酸素状態でも同様の時間経過で同程度のVEGFの誘導が見られた。テトロドトキシン、シクロヘキシミドはそれぞれ虚血感受性、虚血時のVEGFの発現を低下させた。 Study II.リンパ球系株化細胞(U-937細胞)を用いた低酸素条件下の培養で、低酸素刺激開始後1時間をピークとして4時間でコントロールレベルにもどり、さらに低酸素状態に置くと9時間以降のVEGF mRNA発現はコントロールレベル以下となるという時間経過でVEGF mRNAの発現誘導が認められた。PO2=90mmHgのメデイウム中での培養ではVEGF mRNAの誘導をもたらさなかった。さらに外来性アデノシン誘導体:5’-N-ethylcarboxamidoadenosine(NECA)は10-7、10-6Mの濃度で投与後1時間で、濃度依存的にVEGF mRNAの発現を増強した。このNECAの効果はA2-antagonistである3,7-dimethyl-1-propargylxanthine(DMPX)、10-6Mで完全に抑制された。dibutyryl cAMPはVEGF mRNAの発現をNECAと同程度に増強した。さらにAdenosine deaminase、非選択的アデノシン受容体阻害剤:7-(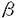 -hydroxyethyl)theophylineおよびアデノシンA2-レセプター阻害剤(DMPX)で低酸素による誘導が抑制された。 -hydroxyethyl)theophylineおよびアデノシンA2-レセプター阻害剤(DMPX)で低酸素による誘導が抑制された。 (考察)Study I.ラットin vivo冠動脈結さつモデル、ランゲンドルフ灌流心および単離培養心筋において虚血あるいは低酸素によりVEGFの発現誘導をもたらすことが明らかになった。虚血心筋におけるVEGF産生細胞は単離培養心筋の結果より心筋細胞である可能性が高いが、後述のVEGFの誘導活性を持つアデノシンのレセプターサブタイプの心組織内分布からすると非筋細胞もその候補者であり、さらに心臓を灌流する血液中の白血球も産生している可能性があると考えられる。テトロドトキシンはエネルギー消費を抑制することで15分程度の虚血時間による心筋内のエネルギーの低下の程度が減少し、結果としてのVEGFの誘導が抑えられた可能性が考えられる。虚血心筋で側副血行路形成促進作用のあるヘパリンにより活性の増加するもうひとつの血管新生因子ファミリーであるbFGFは上述のいずれのモデルにおいてもmRNAの増加は明らかでなかった。VEGFの誘導に必要な虚血の程度は5-10分程度の可逆性の虚血で十分であり、慢性の冠動脈狭窄状態でも間欠的にVEGFが誘導されている可能性が考えられ、VEGFが虚血心における血管新生に関与する可能性を広げるものであると考えられる。 Study II.U-937細胞においてもVEGF mRNAは低酸素刺激により誘導された。低酸素刺激のthresholdはPO2値で40-90mmHgの間にあると考えられた。またアデノシン誘導体であるNECAが濃度依存的にVEGF mRNAを誘導した。低酸素によるVEGF mRNAの誘導がAdenosine deaminase、非選択的アデノシン受容体阻害剤:7-(-hydroxyethyl)theophylineおよびアデノシンA2-レセプター阻害剤(DMPX)で抑制されることは低酸素によるVEGF mRNAの誘導の系に内因性のアデノシンが関与することを示唆するものであると考えられる。ただし、低酸素刺激時の培養メデイウム中のアデノシン濃度は低くenzyme immunoassayの測定感度の下限に入るため低酸素によるアデノシンの増加には測定値上有意差は得られなかった。ランゲンドルフ灌流による心虚血では灌流液中のアデノシン濃度は有意に上昇しており虚血によるアデノシンの細胞外レベルでの増加を裏付けるものであると考えられる。虚血心で増加するアデノシンは心臓に対して陰性変時、変力作用を持ち、また先行する虚血による心筋の虚血耐性獲得の現象への関与が示唆されている。VEGFの虚血組織での働きを考えれば、その発現誘導をもたらすアデノシンは既知の活性以外に、血管新生を通して虚血組織保護に働く可能性があり、一方で悪性腫瘍の治療においてはアデノシン阻害剤は腫瘍栄養血管の進展を抑制することを通してひとつの選択肢になる可能性があると考えられる。 貧血および低酸素で誘導されることの知られているエリスロポエチンとアデノシンの関連は、報告されている限り、低酸素刺激存在下にアデノシンがA2-レセプター作動性に付加的にエリスロポエチンの誘導を増強し、アデノシン受容体阻害剤は低酸素刺激によるエリスロポエチンの誘導を抑制しないというものであり、この点でVEGFとエリスロポエチンの誘導の機構は異なっていることが示唆される。 (結語)3種の心筋虚血モデルで虚血あるいは低酸素によるVEGF mRNAの強い発現誘導がみられた。その時間経過は迅速で1時間前後で発現は最強となった。虚血、低酸素性のVEGF mRNAの誘導について内因性のアデノシンがA2-レセプター作動性に寄与していることが明らかになった。 |