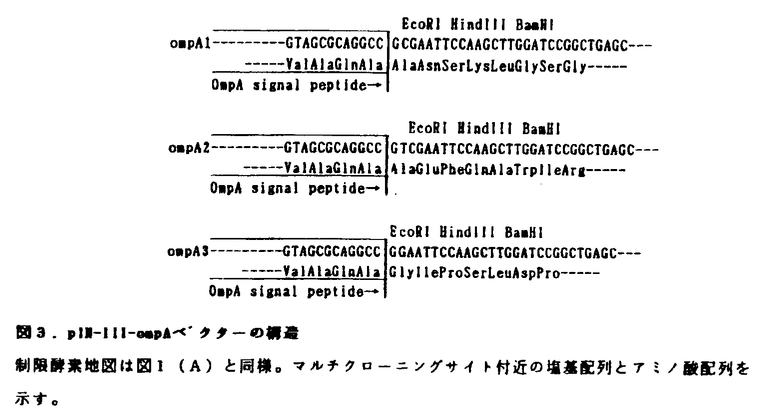
| 審査要旨 | | 本論文は,大腸菌ならびに酵母を宿主として用い,大腸菌の分泌シグナルを利用したタンパク質の部位特異的分泌生産に関する遺伝子工学的研究をまとめたものである。 第1章の序論では,遺伝子工学的なタンパク質の生産方法の中で細胞質内生産を行った場合の諸問題(N末端メチオニンの残留の可能性,ヌクレアーゼやプロテアーゼなどの細胞毒性タンパク質の生産の困難さ等)に触れ,これら問題を克服する方法の一つとして分泌生産が考えられることが述べられている。 第2章では,大腸菌外膜タンパク質であるリポタンパク質(Lpp)の分泌シグナルとStaphylococcus aureusヌクレアーゼAとの融合タンパク質のlpp-lacプロモーター制御下の大腸菌発現プラスミドを作製し,融合部位の違いにより融合タンパク質とそのプロセシング産物の細胞内蓄積部位に相違の見られることを明らかにした。 Lppシグナルの全20アミノ酸配列との融合タンパク質の場合には,シグナルペプチドは切断されず,生産物は細胞質膜画分と一部ペリプラズム画分に見いだされた。細胞質膜に結合した融合タンパク質は,未切断のシグナルペプチドにより細胞質膜に結合し,ペリプラズム側に酵素部分が出ていると考えられた。 LppシグナルとLppのN末端8アミノ酸残基(脂質修飾される1位のシステイン残基を含む)との融合タンパク質の場合には,シグナルペプチドが切断され,脂質修飾された生産物が外膜画分に見いだされた。2時間の発現誘導により,融合タンパク質は全菌体タンパク質の5.7%以上を占める高生産を示した。これらの結果は,Lppシグナルと脂質修飾を用いるこの発現系により,融合タンパク質が外膜に特異的に蓄積することを示している。 第3章では,大腸菌外膜タンパク質A(OmpA)の分泌シグナルを利用したlpp-lacプロモーター新規大腸菌分泌発現用ベクターの作製と,これを用いたヌクレアーゼAならびにヒト・スーパーオキシドジスムターゼ(hSOD)の大腸菌ペリプラズムへの分泌生産について述べている。 外膜タンパク質A遺伝子ompAのシグナルペプチドのコード部分を含み,その3’側にマルチ・クローニング部位(EcoRI,HindIII,BamHIの制限酵素部位)を持ち,任意のコード枠の遺伝子のクローニングが可能となるよう,コード枠のずれた3種類の大腸菌分泌用ベクター(pIN-III-ompA1,ompA2,ompA3)を作製した。 OmpA分泌シグナルーヌクレアーゼA融合タンパク質発現プラスミドpIN-III-ompA-#98を持つ大腸菌では,110分間の発現誘導により全菌体タンパク質の約3%の成熟体がペリプラズムに分泌されることが確認された。 pIN-III-ompA-#98に存在する,シグナルペプチドのC末端とヌクレアーゼAのN末端の間にある余剰配列部分(11アミノ酸コード領域)を部位特異的変異により除去した発現プラスミドpONF1では,同様の誘導条件下で全菌体タンパク質の約10%を占める成熟体がペリプラズムに分泌され,約2%を占める前駆外が細胞質膜に検出された。 細胞質に存在する二量体酵素であるhSODの,OmpAシグナルペプチドとの融合タンパク質の発現プラスミドの場合には,6時間の発現誘導により全菌体タンパク質の約7%の成熟体hSODがペリプラズムに分泌生産されることが確認された。 第4章では,酵母Saccharomyces cerevisiaeを宿主とする,OmpA分泌シグナル-hSOD融合タンパク質のPH05プロモーター制御下の誘導発現系を作製し,hSODの分泌生産について検討している。その結果,発現プラスミド保持酵母の低リン酸あるいは無リン酸誘導培地での培養により,培地中に細胞質性タンパク質hSODか成熟型で分泌されることを確認した。 以上,本研究は,大腸菌の新規分泌発現ベクターの作製を行い,大腸菌のシグナルペプチドを利用した,大腸菌ならびに酵母を宿主とする異種タンパク質の部位特異的分泌生産について明らかにしたものであり,学術上,応用上貢献するところが少なくない。よって審査委員一同は申請者に博士(農学)の学位を授与してしかるべきものと判定した。 |