本研究は動物体内での窒素化合物の代謝の最終段階を明らかにするために、亜硝酸レダクターゼのうち銅酵素を精製し、シトクローム酵素との関係、ことに電子供与体としての特性を明らかにする目的で行われた。Achromobacter cycloclastesの亜硝酸レダクターゼの銅酵素を精製し、分析したところ下記の結果を得た。 1.吸光度特性: Achromobacter cycloclastesの青銅タンパク質を完全に精製したところ、最大吸収波長が750、595、453及び278nmを示し、還元すると278nm以外の吸収は消失した。亜硝酸レダクターゼを完全に精製したところ、可視部での吸収は695、585及び458nmであり、還元するとこれらの吸収は消失した。2種類のシトクローム(C551及びC554)は弱アルカリ性のタンパク質であって、hydroxylapatiteカラムで両者を分離し、さらにCM-Biogel及びSephadexカラム等によって両者を完全に精製することができた。C551及びC554はアスコルビン酸で完全に還元された。C551は酸素存在下の吸収が524、409及び280nmを示した。還元状態では551、521.5及び415nmであった。C554は酸素存在下の吸収が521、412及び280nmであった。還元状態では554.5、523及び418nmであった。 2.分子量及びアミノ酸配列: 青銅タンパク質の分子量は11,800であり、銅はタンパク質1分子あたり0.91分子含まれていた。アミノ酸組成及びN末端アミノ酸配列も決定し、Alicaligenes faecalis S-6から精製した青銅タンパク質と比較している。亜硝酸レダクターゼの分子量は69,000で、タンパク質1分子あたり銅の含有量は3分子であった。アミノ酸組成及びN末端アミノ酸配列も決定している。この酵素の比活性は376,000mmole/mg/hrにも達し、これまでの報告値よりも2倍ほど高かった。シトクロームC551およびC554の分子量は17,000であった。C551は2個、C554は1個のC型ヘムを持っていることがわかった。 3.EPRの特性:青銅タンパク質は77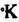 の条件下では0.5spin、36では0.48spinであり、この値は典型的なI型Cu2+の存在を示し、II型Cu2+は含まれていなかった。亜硝酸レダクターゼは77では3.1spin、36では2.6spinであり、73Gにhyperfinesplittingが見られた。しかし、168Gに幅広いピークが見られ、I型及びII型のCu2・を同時に持つことを示した。これをアスコルビン酸で還元すると、I型Cu2+のみが影響された。還元すると全てのEPR signalが消失し、亜硝酸を加えると全てのsignalが再び現れたので、亜硝酸はこの酵素に対して影響及ぼすものと考えられた。またEPR差スペクトルの手法を用いると、亜硝酸はII型Cu2+に対すして影響を及ぼすことが示された。この酵素をsodium dithioniteで滴定をしたところ、I型Cu2+の酸化還元電位は+243mVであり、II型Cu2+は約+83mVであった。 の条件下では0.5spin、36では0.48spinであり、この値は典型的なI型Cu2+の存在を示し、II型Cu2+は含まれていなかった。亜硝酸レダクターゼは77では3.1spin、36では2.6spinであり、73Gにhyperfinesplittingが見られた。しかし、168Gに幅広いピークが見られ、I型及びII型のCu2・を同時に持つことを示した。これをアスコルビン酸で還元すると、I型Cu2+のみが影響された。還元すると全てのEPR signalが消失し、亜硝酸を加えると全てのsignalが再び現れたので、亜硝酸はこの酵素に対して影響及ぼすものと考えられた。またEPR差スペクトルの手法を用いると、亜硝酸はII型Cu2+に対すして影響を及ぼすことが示された。この酵素をsodium dithioniteで滴定をしたところ、I型Cu2+の酸化還元電位は+243mVであり、II型Cu2+は約+83mVであった。 ferricyanideで酸化すると、この還元性酵素はII型Cu2+、I型Cu2+が還元状態であることがわかった。青銅タンパク質もsodium dithioniteで滴定すると酸化還元電位は約+308mVでることがわかるが、亜硝酸は青銅タンパク質とは反応しなかった。 4.亜硝酸レダクターゼの電子供給体としての青銅タンパク質:青銅タンパク質を還元した後、酸化型亜硝酸レダクターゼを加えると595nmの吸収が現れることから、青銅タンパク質が酸化されたことが示唆された。同時に亜硝酸レダクターゼが還元される。 5.亜硝酸レダクターゼとシトクロームの相互作用:Achromobacter cycloclastesのシトクロームは、C551及びC554ともに亜硝酸レダクターゼの電子供給体となることはできなかった。 青銅タンパク質のみが亜硝酸レダクターゼの電子供給体となることができ、同じ細菌から分離したシトクロームC551、C554はいずれも電子供給体・電子受容体となることができなかったことは、この細菌では青銅タンパク質が亜硝酸レダクターゼの生理的な唯一の電子供給体であることを示したものである。EPRの解析は青銅タンパク質が1個のI型Cu2+を含んでいることを示し、一方、亜硝酸レダクターゼはI型とII型のCu2+を含んでいて、青銅タンパク質を還元できることを明らかにした。さらに、Stopped-flowの研究により、2つ異なるタンパク質の間の作用、反応速度及び効率に対する解釈がはっきりとしてきた。また、亜硝酸がこの2つのタンパク質の間の反応に促進効果があることが示された。 以上、本研究は亜硝酸レダクターゼのうち銅酵素が電子を供給していて、シトクロームは電子供与体にならないことを生化学的、物理化学的手法により明らかにしたものであるが、窒素代謝におけるNOを反応産物とする亜硝酸レダクターゼの生理的役割を理解する上で重要な反応機構を明らかにしたものとして学位の授与に値するものと考えられる。 |