学位論文要旨
| No | 212515 | |
| 著者(漢字) | 杉田,繁夫 | |
| 著者(英字) | Sugita,Shigeo | |
| 著者(カナ) | スギタ,シゲオ | |
| 標題(和) | H1N1型ブタ及びヒトインフルエンザAウイルスの血球凝集素遺伝子の分子進化 | |
| 標題(洋) | Molecular Evolution of Hemagglutinin Genes of H1N1 Swine and Human Influenza A Viruses | |
| 報告番号 | 212515 | |
| 報告番号 | 乙12515 | |
| 学位授与日 | 1995.10.25 | |
| 学位種別 | 論文博士 | |
| 学位種類 | 博士(医学) | |
| 学位記番号 | 第12515号 | |
| 研究科 | ||
| 専攻 | ||
| 論文審査委員 | ||
| 内容要旨 | 1918年に2,000万人以上の死者を出したスペイン風邪ウイルスは、H1N1型インフルエンザウイルスと推定され、ヒト世界では抗原性を急速に変化させ現在のロシア(ソ連)風邪ウイルスとして流行を繰り返しているが、ブタ世界においては、スペイン風邪ウイルスの抗原性を保持したままブタの中で流行を繰り返し現在に至っていると考えられている(図1)。 しかし、1976年に米軍内で流行し、抗原解析の成績よりブタ型インフルエンザウイルスとされているヒトからの分離ウイルスであるA/New Jersey/11/76を除いて、抗原性を担っている血球凝集素(HA)タンパクをコードするブタインフルエンザウイルス野外分離株のHIHA遺伝子配列の報告は全くない。そこで、1930年に初めてShopeがクローニングに成功したスペイン風邪ウイルスと密接な関係があると推定されているブタ・インフルエンザウイルスA/sw/Iowa/15/30(IOWA30)をはじめ、ブタ・インフルエンザウイルスであるA/sw/Illinois/63(ILL63)、A/sw/Hong Kong/74(HK74)及びA/sw/Ehime/1/80(EHM80)のHA遺伝子をクローニングし、遺伝子配列を決定することにより、ヒト・インフルエンザウイルスとブタ・インフルエンザウイルスの分子進化学的関係を解析した。 クローニングし決定したブタ・インフルエンザウイルスの遺伝子配列とこれまでに報告された遺伝子配列を基に、同義座位について全ての株間の組み合わせについて遺伝子距離を計算し、近隣接合法によって系統樹を作成した(図2)。同義置換は、アミノ酸配列の変化を伴わない変異なので、同義座位でウイルス変異速度を計算するとHAタンパクの変異を伴うことによる自然淘汰の影響を考える必要がなく、RNAボリメラーゼの読み間違いによるランダムな変異速度を反映するものと考えられている。 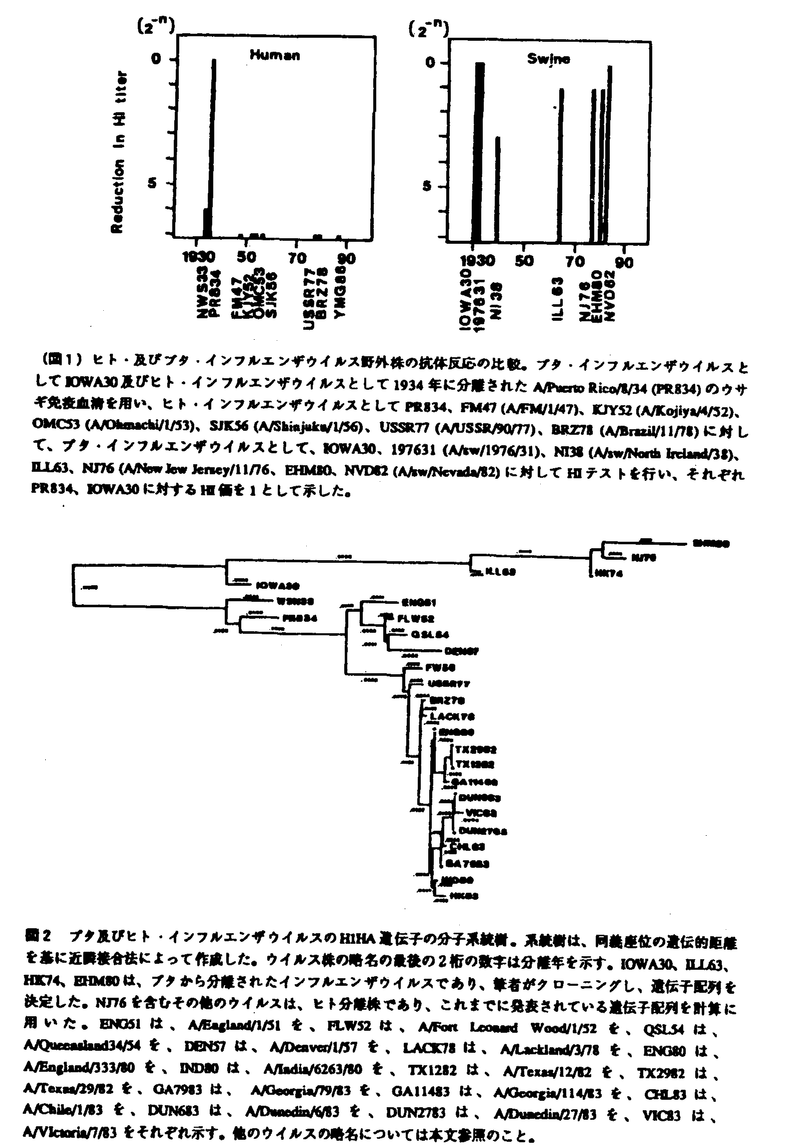 系統樹では、IOWA30、ILL63、HK74、EHM80のブタ・インフルエンザウイルス及びブタ型インフルエンザウイルスと考えられているNJ76が1つのグループを形成し、その他のヒト型インフルエンザウイルスのグループとは違っていた。また、ブタ、ヒト型インフルエンザウイルスともに、年代にしたがって新しいものはど右に描かれ、年代に従って遺伝し変異が蓄積し進化していることが示唆されるが、ヒト型インフルエンザウイルスの糸統樹上FW50(A/Fort Warren/1/50)とUSSR77は、非常に近接しており、27年の分離年の違いが系統樹に反映されておらず、USSR77の進化が、およそ25年間凍結していたと考えられる。 進化速度をさらに正確に求めるために、ブタ型インフルエンザウイルスについては、IOWA30と他のブタ型インフルエンザウイルスを系統樹上の分岐点を仮想原点とし、また、ヒト型インフルエンザウイルスの場合は、WSN33と他のヒト型インフルエンザウイルスの系統樹上の分岐点を仮想原点とし、仮想原点からの系統樹上の遺伝的距離を分離年に対してプロットした(図3)。すると、ブタ、ヒト型インフルエンザウイルスともに非常に良い直線関係が得られたが、ヒト型インフルエンザウイルスの場合、ロシア風邪ウイルス(B)とそれ以外の古典的H1N1ウイルス(A)の2つの直線が得られ、FW50とUSSR77の間に25年間のHA遺伝子の進化の凍結現象が観測された。 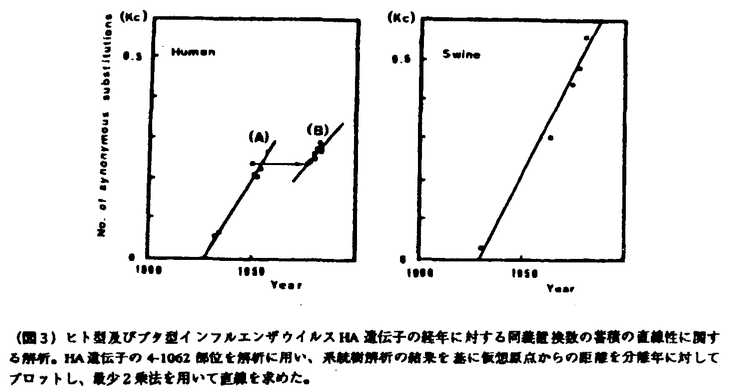 さらに、図3の直線の傾きよりインフルエンザウイルスの進化速度を求めたところ、同義座位での進化速度は、ヒトインフルエンザもブタインフルエンザもほぼ同じであった(表1)。また、同様の手法で非同義座位での進化速度を求めたところ、ブタインフルエンザのそれは、ヒトインフルエンザの約1/3であった。ブタインフルエンザウイルスの抗原性変異が遅いのは、アミノ酸変異を伴う遺伝子変異が制限されているためと思われる。 非同義置換(アミノ酸配列の変化を伴う遺伝子変異)は、その変異によってもたらされるアミノ酸配列の変化がウイルスの生存にとって不利である場合は、自然淘汰により変異ウイルスは自然界から除去されるが、非同義座位も同義座位も同じ確率でポリメラーゼにより遺伝子の読み間違いは起こっている。つまり、非同義座位での変異の確立を同義座位での変異の確立に対して比較すると、自然淘汰によるアミノ酸配列の変化による影響を調べることができる。そこで、PR8株で決められた30の抗原決定基のアミノ酸配列について同義座位の変異と非同義座位の変異の割合を解析すると、ヒト・インフルエンザウイルスは、ほぼ同じ様な割合で起こっているが、ブタ・インフルエンザウイルスでは非同義置換の割合が少なく、抗原決定基であってもブタ・インフルエンザウイルスの場合、変異が強く制限されておりブタ・インフルエンザウイルスのアミノ酸配列の変異がタンパクの機能上制約されていることがわかった(図4)。ブタ・インフルエンザウイルスの抗原決定基がヒト・インフルエンザウイルスと同じサイトにあると仮定すると、ブタ・インフルエンザウイルスが抗原性を保ったまま変異進化している理由は、タンパクの機能的制約が強いためスペイン風邪ウイルスの機能を保持したまま進化し、僅かな復帰突然変異でスペイン風邪で見られたようなヒトに対する大きな感染性、重篤な病原性を獲得する可能性があることを示唆している。実際に、1976年にブタ・インフルエンザウイルスがヒトに感染し米軍内で流行して以来、ヒトに対して時々感染を起こしていること、1988年には、アメリカでブタ型インフルエンザによって32歳の妊婦が死亡するなどの死亡症例も報告されており、ブタ・インフルエンザウイルスは、僅かの突然変異によってヒト世界で大流行し、重篤な病原性を獲得し、甚大な被害をもたらす可能性がある要注意ウイルスであると考えられる。ただし、上記の推定は、ブタ・インフルエンザウイルスとヒト・インフルエンザウイルスの抗原決定基が同一部位にあるという仮定に基づくものであるので、今後、ブタ・インフルエンザウイルスの抗原決定基を明らかにし、同様の比較を行う必要がある。 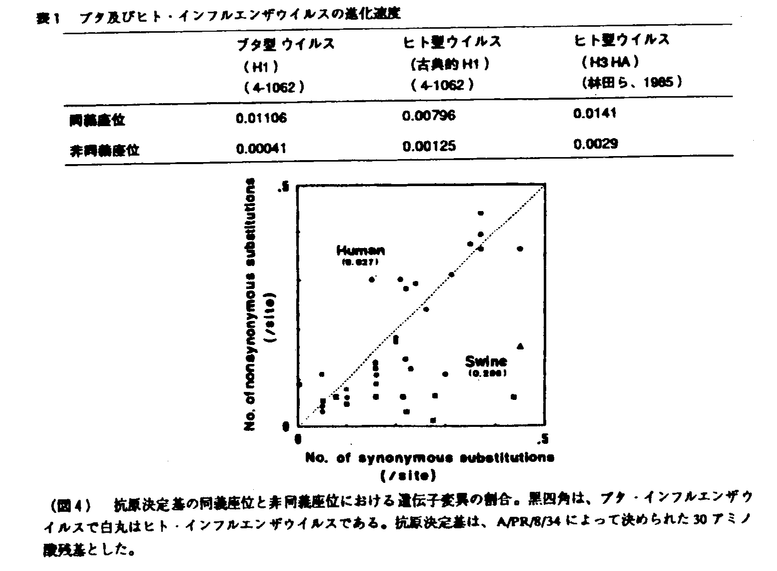 今回のブタ・インフルエンザウイルスの解析に用いることのできたデータは非常に限られたものであった。これは、世界中からH1N1型野生ブタ・インフルエンザウイルス株を集めることが困難であったためである。また、野生株を培養細胞株または鶏卵を用いてクローニングしたために、野生流行株の遺伝子配列を直接決定することができなかった。これらのことから、今回の解析結果がブタ・インフルエンザウイルスの進化そのものを反映しているかどうかには疑問が残る。RNAウイルスゲノムの変異が非常に速いことを考えると、今後の課題として、これらの問題点を踏まえたさらなる研究の努力が必要である。 | |
| 審査要旨 | 本研究はインフルエンザAウイルスの中和において最も重要な役割を担っている血球凝集素タンパクの抗原変異のメカニズムを明らかにするため、変異が遅いとされているブタインフルエンザウイルスと変異が速いとされているヒトインフルエンザウイルスの変異パターンの違いを分子進化学的に解析することにより、インフルエンザAウイルスの血球凝集素タンパクの変異のメカニズムを明らかにしようとしたものであり、下記の結果を得ている。 1.世界中で2,000万人以上が死亡したスペイン風邪の原因と考えられているインフルエンザウイルスに抗原性が最も近いと考えられているA/sw/Iowa/15/30(IOWA30)をはじめ、A/sw/Illinois/63(ILL63)、A/sw/Hong Kong/74(HK74)及びA/sw/Ehime/1/80(EHM80)の血球凝集素遺伝子が世界で初めてクローニングされ、血球凝集素遺伝子の全塩基配列が示された。H1型ブタインフルエンザウイルスの血球凝集素遺伝子の配列報告は本報告が最初である。 2.決定されたブタインフルエンザウイルスの遺伝子配列とこれまでに報告されたいるヒトインフルエンザウイルスの遺伝子配列を基に、系統樹が描かれた。系統樹では、IOWA30、ILL63、HK74、EHM80のブタインフルエンザウイルス及びブタ由来ヒト・インフルエンザウイルスと考えられているNJ76は、ヒトインフルエンザウイルスと異なる1つのグループを形成し、2つのグループが分岐したのは、1918年のスペイン風邪ウイルスの大流行より以前の1905頃であることが示された。 3.決定された遺伝子配列を基にインフルエンザウイルスの進化速度が解析され、同義座位での進化速度は、ヒトインフルエンザウイルスもブタインフルエンザウイルスもほぼ同じであること、非同義座位での進化速度は、ブタインフルエンザウイルスがヒトインフルエンザの約1/3であることが示された。 4.抗原決定基のアミノ酸配列について同義座位と非同義座位の変異の割合が解析され、ヒトインフルエンザウイルスではほぼ同じ割合で起こっているのに対し、ブタインフルエンザウイルスでは非同義置換の割合が少なく、アミノ酸変異が強く制限されていることが示された。 5.以上の知見から、ブタインフルエンザウイルスが抗原性を保ったまま変異進化している理由は血球凝集素タンパクのアミノ酸配列の変異に対する機能的制約が強いためであることが示唆された。つまり、現在のH1N1ブタインフルエンザウイルスは、スペイン風邪ウイルスの機能を保持したまま進化し、僅かな復帰突然変異で1918年のスペイン風邪で見られたようなヒトに対する大きな感染性、重篤な病原性を獲得する可能性があることが示唆された。 以上、本論文はH1N1型ブタインフルエンザウイルスの血球凝集素遺伝子配列を初めて決定し、ヒトインフルエンザウイルスとの分子進化像の違いを明らかにした。解析に用いることのできたデータ数が少ないため、ブタインフルエンザウイルスとヒトインフルエンザウイルスの違いを明快に示したとは言えないものの、非同義置換がブタインフルエンザウイルスで強く抑えられていることを解明した点はインフルエンザウイルスの進化像を知る上で重要な貢献をなすと考えられ、学位の授与に値するものと考えられる。 | |
| UTokyo Repositoryリンク | http://hdl.handle.net/2261/50671 |