| 内容要旨 | | fyn遺伝子はsrc遺伝子群に属する癌関連遺伝子であり,分子量約6万の非受容体型チロシンキナーゼをコードしている。その遺伝子産物Fynは,主としてリンパ系組織ならびに中枢神経系における発現の高いことが知られており,T細胞においてはT細胞抗原受容体複合体と物理的に緩やかに会合している。また,Srcファミリーに属する他のメンバーであるLckは,CD4またはCD8と会合している。このようなことから,Fynを含むSrcファミリーチロシンキナーゼが,さまざまな細胞の活性化シグナル伝達系で重要な役割を果たしていることが示唆されている。Fynは細胞膜に結合しうるアミノ末端を含むFynに特異的な領域と,Srcファミリー間で類似性の高い調節領域(SH2およびSH3)とキナーゼ領域から構成されている。キナーゼ領域の相違により,胸腺型(T-Fyn)と脳型(B-Fyn)に区別することができる。 一方,HIV遺伝子の発現は,T細胞の活性化にともなって上昇することが知られている。そこで,FynがHIV遺伝子プロモーターを介した転写の活性化に関与している可能性を検討するために,Fyn発現プラスミドをT細胞株Jurkatに導入,発現させ,HIVLTRからの転写におよぼすFynの影響を調べた。Fyn発現ベクターとして,野生型のT-fyn,さらに528番のチロシンをフェニルアラニンに置換した高いチロシンキナーゼ活性を示す活性型T-fynF,SH2領域の一部を欠失させたt-1を用い,HIVLTRをCAT遺伝子につないだHIV-CATによるCAT活性を測定した。なお,CAT活性の差を見やすくするために,T細胞を活性化するConA,およびPKCを活性化するPMAによる刺激を加えた。 CATアッセイの結果,野生型Fynを過剰発現させることにより,ConAおよびPMAの刺激にともなうHIVプロモーターからの転写が亢進することが示された。また,活性型Fynを用いると野生型FynよりもさらにCAT活性が上昇した。Lckを導入した場合にも,野生型および活性型Lckの両者において,Fynより弱いながらも同様の結果が得られた。一方,Srcファミリーに属する他のメンバーであるLynを導入した場合,野生型ならびに活性型LynのいずれにおいてもCAT活性の上昇をみなかった。これは,LynがT細胞で発現していないことに関連すると考えられる。以上から,FynとLckが特異的にHIVLTRからの転写活性を上昇させることが結論された。 次に,FynによるHIV-CATの活性化に必要なHIVLTR部位を詳細に検討するために,HIV-CAT変異体を用いてCATアッセイを行なった。HIVLTRに作用してウイルスの転写を活性化する細胞側の因子として,NF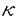 B,NFAT1,Sp1,LBP1などの転写因子が知られているが,ここではNFBに注目した。プロモーター領域を5’側から順次欠失したHIV-CAT変異体CD7,CD16,CD23まではNFB結合部位を保有しているが,次のCD52ではNFB結合部位が欠失している。E5とE8はCD52の5’側に合成したB結合部位を導入したものである。さらに,B結合部位のみにポイントミューテーションを入れたBmutantをあわせ,合計7種類のHIV-CAT変異体を用意した。 B,NFAT1,Sp1,LBP1などの転写因子が知られているが,ここではNFBに注目した。プロモーター領域を5’側から順次欠失したHIV-CAT変異体CD7,CD16,CD23まではNFB結合部位を保有しているが,次のCD52ではNFB結合部位が欠失している。E5とE8はCD52の5’側に合成したB結合部位を導入したものである。さらに,B結合部位のみにポイントミューテーションを入れたBmutantをあわせ,合計7種類のHIV-CAT変異体を用意した。 CATアッセイの結果,B結合部位を越えてHIVLTRを欠失させると,ベクターのみを導入した場合と活性型Fynを導入した場合の間でCAT活性に差がなくなった。さらに,B結合部位を越えて欠失させたレポータープラスミドCD52と,これに合成したB結合部位を導入したレポータープラスミドE5とE8の間でCAT活性の差を調べた。その結果,CD52ではFynによるCAT活性の上昇がみられなかったが,E5では約6倍,E8では約3倍のCAT活性の上昇が認められ,その活性は野生型HIV-CATによるものと同程度であった。このことから,FynによるHIV-CATの活性上昇には,少なくともNFB結合部位が関与していることが明らかにされた。しかしながら,B結合部位のみにポイントミューテーションを入れたBmutantでは,FynによるCAT活性の弱い上昇が認められた。この原因として,B結合部位以外にCAT活性上昇に関与する部位の存在が考えられる。 なお,FynからSH2領域の一部を欠失させたt-1を用いると,活性型Fynによって上昇したCAT活性が低くなった。このことは,Fynからのシグナルを下流に伝達するうえで,FynのSH2領域が重要であることを示唆している。 ここで,Fynからのシグナル伝達の下流に,細胞質内NFBを活性化するPKCの存在を考慮し,PKCのインヒビターであるH7の存在下でCATアッセイを行なった。その結果,H7を加えることによりFynによるHIV-CATの活性上昇が一部阻害されることがわかった。このことは,FynからのシグナルがHIVLTRに伝達される過程でPKCの作用を一部介していることを示しており,Fynを介したNFBによるHIV-CATの活性上昇が考えられた。 さらに,FynによるHIVLTRを介したCAT活性の上昇が,NFBを含み,Fynにより調節されるB配列に結合する転写活性化因子によって媒介されている可能性を検討した。まず,T細胞ハイプリドーマHBC21.7.31を用いて活性型B-Fynの高発現細胞154を樹立し,これらの細胞をanti-CD3刺激後,核内タンパク質を調製してゲルシフト法による解析を行なった。 HIVLTRのB部位をプローブに用いた実験では,B1からB4までの4種類のバンドが検出できた。これらのバンドの特異性は,HIVのB部位とその変異体,さらにIL2R とMHC-IのB部位を合成,これらをコンペティターとしたコンペティション実験で検証した。その結果,B1とB2では野生型HIV,IL2RとMHC-IのB部位をコンペティターとしてプローブの50倍量加えることによってバンドが消失した。このことは,B1とB2が特異的にB部位と結合することを示している。一方,B3とB4のバンドは,野生型HIVのBコンペティターでのみバンドが消失した。このことは,B3とB4がHIVのB部位に特異的に結合することを意味しているが,さらなる解析が待たれる。 とMHC-IのB部位を合成,これらをコンペティターとしたコンペティション実験で検証した。その結果,B1とB2では野生型HIV,IL2RとMHC-IのB部位をコンペティターとしてプローブの50倍量加えることによってバンドが消失した。このことは,B1とB2が特異的にB部位と結合することを示している。一方,B3とB4のバンドは,野生型HIVのBコンペティターでのみバンドが消失した。このことは,B3とB4がHIVのB部位に特異的に結合することを意味しているが,さらなる解析が待たれる。 また,Fyn高発現株とその親株との間で,これらのバンドへの影響を調べた。c-erbB-2プロモーター部位をプローブとして用いたコントロール実験では,Fyn高発現細胞とその親株の間でシフトしたバンドの強さに違いはなかった。しかし,B部位をプローブにした実験では,B1とB2においてFyn高発現細胞のほうがより強いバンドがみられた。これに対して,B3とB4のバンドでは,有意な差は認められなかった。このことから,FynからのB部位へのシグナルが作用するうえでB1とB2を構成するタンパク質がその役割を担っていることが示唆された。 さらに,B1とB2を構成しているタンパク質の大きさを決定するために,UVクロスリンク法を用いた。チミンをプロモデオキシウリジンに置換したBプローブを用い,UVで架橋されたBプローブとタンパク質の結合物をゲルシフト後にゲルから切り出し,エレクトロエリューションにより回収,SDS-PAGEで解析を行なった。その結果,B1は100,65,60と50kd,B2は65と50kdのタンパク質から構成されることが明かになった。また,NFB/c-Relに交叉反応するモノクローナル抗体を用意し,抗体と反応させた後Bプローブと反応させてゲルシフトを行なうとB1とB2のバンドが薄くなった。このことにより,B1とB2が65および50kdのNFBを含んでいることが示された。一方,B3とB4は,それぞれ56と45kdのタンパク質であった。 さらに,抗CD3抗体の刺激によるB結合タンパク質の細胞質と核内の分布パターンをゲルシフト法により解析した。その結果,刺激前には細胞質内に豊富に存在するB1とB2が,抗CD3抗体の刺激により核内に移行するが,その移行が活性型T-Fynの高発現細胞では親株に比べてさらに促進された。これは,NFBはIBと結合して細胞質内に存在するが,PKCによるIBのリン酸化により核内へNFBが移行するモデルに類似している。実際FynからのHIVLTR転写活性化にIBを不活化するPKCが関与していることは,H7を使ったCATアッセイで示したとおりである。 以上の実験結果をまとめると,次のことが結論された。(I)FynからのシグナルがHIVLTRの転写活性を上昇させる。(II)Fynからのシグナル伝達にはFynのSH2領域が重要である。(III)HIVLTR上のB配列がFynからの転写制御に関与している。(IV)Fynの高発現細胞では核内に存在するB配列結合タンパク質量が上昇する。(V)抗CD3抗体の刺激後,B配列結合タンパク質の細胞質から核内への移行がFynの高発現により増強する。(VI)B配列結合タンパク質にはNFBが含まれている。以上より,Fyn遺伝子産物がPKCを介したHIVプロモーターの転写制御に関与していることが示唆された。 |