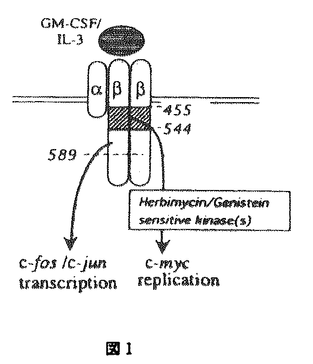
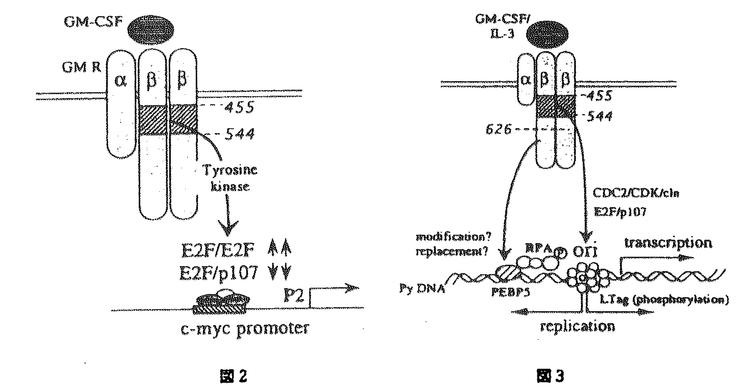
本研究は様々な血球系細胞の増殖と分化に重要な役割を果たしているGM-CSFのレセプターのシグナル伝達の機構を検討するために、血球系および非血球系の培養細胞株に野生型または変異体のヒトGM-CSFレセプターを再構成し種々のシグナル伝達について検討を加えたものである。この研究によって得られた主な結果は以下のようなものである。 1.GM-CSFレセプターは、チロシンキナーゼレセプターやホルモン型レセプターなどこれまで知られたレセプターとは構造を異にするサイトカインレセプターファミリーに属し、そのシグナル伝達は血球系に特異的である可能性が指摘されていた。本研究により線維芽細胞に再構成されたGM-CSFレセプターがDNA合成、初期応答遺伝子の転写活性化などのシグナルを伝えうることが示され、血球系以外の細胞でもシグナル伝達が可能であることが明らかになった。また、c-fosプロモーターを用いた一過性発現の系によるレセプターシグナル伝達の評価が可能になり、レセプターシグナル伝達の研究がより迅速に正確に行なうことが可能になった。 2.前述のようにGM-CSFはレセプター自身、細胞内蛋白質のチロシン残基のリン酸化に加えて、増殖、c-fos、c-jun、c-mycなどの初期応答遺伝子の活性化を血球糸細胞、非血球系細胞でおこすが、これらの活性化にかかわるシグナル伝達分子、チロシンキナーゼの関与について不明の点が多かった。GM-CSFレセプター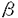 鎖の段階的欠失変異体、チロシンキナーゼインヒビターを用いて、これらの活性を制御するシグナル伝達経路が大きく2つの経路に大別されることを明らかにした。第1の経路はc-mycの活性化、増殖に至るもので、チロシンキナーゼインヒビター、ゲニステインによって完全に阻害され、ヒトGM-CSFレセプター鎖の膜貫通領域直下部分のみを必要とする。第2の経路はc-fos、c-junの活性化に至りヒトGM-CSFレセプター鎖膜貫通直下部分に加えてよりC末端部分も必要とする。またこの経路はゲニスタインによって全く阻害されないばかりか、c-fosの活性化はより増強されることが明らかになった。これらの結果によりチロシンキナーゼが実際にGM-CSFのシグナル伝達に関与していること、またレセプター変異体の利用により特定のシグナル経路のみを活性化または不活性化することができるようになり、レセプターシグナルの解析に新しい展開を与えた。 鎖の段階的欠失変異体、チロシンキナーゼインヒビターを用いて、これらの活性を制御するシグナル伝達経路が大きく2つの経路に大別されることを明らかにした。第1の経路はc-mycの活性化、増殖に至るもので、チロシンキナーゼインヒビター、ゲニステインによって完全に阻害され、ヒトGM-CSFレセプター鎖の膜貫通領域直下部分のみを必要とする。第2の経路はc-fos、c-junの活性化に至りヒトGM-CSFレセプター鎖膜貫通直下部分に加えてよりC末端部分も必要とする。またこの経路はゲニスタインによって全く阻害されないばかりか、c-fosの活性化はより増強されることが明らかになった。これらの結果によりチロシンキナーゼが実際にGM-CSFのシグナル伝達に関与していること、またレセプター変異体の利用により特定のシグナル経路のみを活性化または不活性化することができるようになり、レセプターシグナルの解析に新しい展開を与えた。 3.c-fos、c-junの活性化に至るシグナル伝達経路がMAPKカスケードの経路の関与などチロシンキナーゼ型レセプターの研究を通じて明らかであるのに対して、c-mycの活性化、増殖の活性化に至るシグナル伝達は未知の部分が多い。この点を解明するためまずGM-CSFによるc-mycの転写活性化機構を明らかにする目的でc-mycプロモーターを用いた増殖因子依存性の一過性発現の系を血球系細胞において確立した。c-mycプロモーターは転写活性が弱くこれまで増殖因子に依存したプロモーター活性の評価系は確立していなかったのでこの解析系が、比較的遺伝子導入効率が悪いとされている血球系細胞で確立されたことは意義が深い。本研究では種々のc-mycプロモーターフラグメントをCAT遺伝子につなぎGM-CSFによる転写活性化に必要な領域を決定し、さらにc-mycプロモーターフラグメントDNAを用いたゲルシフト解析による転写因子の結合状態の解析によりGM-CSFは主にc-mycプロモーターのP2転写開始部分を使っていること、E2F部位を介して正、負の調節を行なっていることなどが明らかになった。これらの事実が明らかになったことにより、さらにGM-CSFにより活性化を受けるE2Fの同定、その制御シグナルの解析へと発展させることが可能になり、これまで未知であったc-myc活性化の機構の解析の解明への道が開けた。 4.さらに細胞増殖の制御機構について明らかにするためにDNA合成の開始機構を解析するためポリオーマウイルスの複製オリジンをもちいたDNA合成開始の解析を開発した。従来、DNA合成、細胞増殖の解析は細胞数の増加、MTT解析、[3H]Thymidineのとりこみなどが用いられているが、これらは細胞の活性を総合して評価しているものであったり、DNA合成の開始、修復、延長反応の総和であるなど特定の反応の解析ではないのでシグナル伝達機構の解析には不向きであった。哺乳細胞ではDNA合成の開始点は同定されておらず、複製装置をへリカーゼ以外全てホストの細胞によっているポリオーマウイルスはよいモデルになると考えられる。本研究で使われたポリオーマウイルスレプリコンは種々のエンハンサーに加えてCAT遺伝子を組み込んであるので転写と複製両者について開始機構の解析が可能である。この系の確立によってGM-CSFが異なるシグナル伝達経路を介してポリオーマウイルスレブリコンの転写と複製を活性化すること、その際エンハンサーが異なる機構で転写と複製に寄与する可能性が示唆された。 以上、本論文はGM-CSFレセプターのシグナル伝達の機構を明らかにし、サイトカインレセプターシグナル伝達全般の解明にも重要な貢献をもたらすと考えらる。さらには未知の部分が多い哺乳細胞でのDNA合成開始の機構解析の格好のモデル系としての有用性も示され、学位の授与に値するものと考えられる。 |  鎖と全てに共有される
鎖と全てに共有される