無脊椎動物の視細胞において細胞内Ca2+は明順応、暗順応に重要な役割を果たしている。無脊椎動物における視細胞の機能についてはカブトガニ(Limulus)による研究がよく知られている。一方、フジツボ(Balanus)の視細胞では、カブトガニと異なり、光刺激によりCaチャンネルを介してCa2+流入を生じることが判明しているので、光刺激による細胞内Ca2+濃度([Ca2+]1)の上昇が大きく、より効率的なCa2+排出機構が発達している可能性が推定される。またフジツボの巨大視細胞は、直径100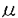 mに及び、膜電位と[Ca2+]1の同時測定が比較的容易である。本研究ではフジツボの巨大視細胞を用い、細胞内Ca2+の恒常性メカニズムについて検討した。その結果、Na-Ca交換輸送によるCa2+のくみ出しが重要であること、また更に、Ca-H交換輸送の関与の可能性が示唆された。 mに及び、膜電位と[Ca2+]1の同時測定が比較的容易である。本研究ではフジツボの巨大視細胞を用い、細胞内Ca2+の恒常性メカニズムについて検討した。その結果、Na-Ca交換輸送によるCa2+のくみ出しが重要であること、また更に、Ca-H交換輸送の関与の可能性が示唆された。 まずフジツボ巨大視細胞に光刺激を与えて、膜コンダクタンスと膜電位に対する影響を検討した(図1)。コントロール溶液(NBS.)(in mM;NaCl 462,KCl 8,CaCl2 20,MgCl2 12,Hepes 10,pH=7.4)中で光刺激を与えると(A)、急速な脱分極が生じ(上段)、それに伴って、一定の電流パルス(下段)に対する膜電位の変化、即ち膜コンダクタンスが上昇した。脱分極及びコンダクタンスは、光刺激中に徐々に回復し、光を遮断すると、急速に回復し、膜電位は過分極方向にリバウンドした(post illumination hyper polarization)。このリバウンド現象は、ウワバインでブロックされることからNa-K ATPaseによる能動輸送を介する現象と報告されているが、実際、明らかなコンダコタンス変化を伴わず、細胞外液からK+を取り除きNa+をLi+に交換する(0-K,0-Na(Li))と消失した(B)。この現象はカブトガニでは報告されていない。 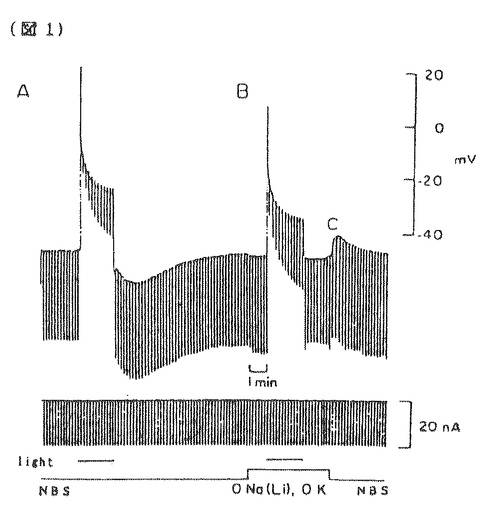 図1 図1 0-Na(Li)液中においては、光刺激によりNBS中と同様に脱分極を生じた(B)。従ってこの脱分極は、Li+が、Naチャンネルを通過することによると考えられる。光刺激中における0-Na(Li)液中でのコンダクタンスの減少は、NBS中より急速であった。この理由の一つの可能性は、0-Naでは、逆向きNa-Ca交換輸送のため、[Ca2+]1が上昇していることである。光刺激遮断後に、0-K,0-Na液よりNBSに戻したところ、一過性の脱分極が起こった(C)。この脱分極は、[Ca2+]1が上昇したために、Na-Ca交換輸送が活性化されたことによると考えられる。 次に明順応からの光感受性の回復を検討した。10秒毎の0.5秒間に光パルス刺激を与え、これにより生じる膜電位変化を測定し視細胞の光感受性の指標とした(図2)。次に持続的光刺激を与えた。膜電位は、一旦脱分極後ゆるやかに回復する明順応応答が認められた。光を遮断すると一過性過分極が生じたが、この時点で、視細胞の光感受性の低下が認められた。膜電位が回復する時間経過とほぼ平行して、光感受性の回復が認められた。次に同様の実験を0-Na(Li)液中で行ったところ、より急速かつ大きな明順応が生じた。この事実はNa-Ca交換輸送の抑制によって、細胞内Ca2+の排出が妨げられ、これによって明順応が促進されたと解釈される。 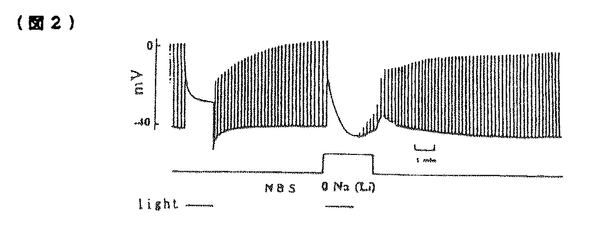 図2 図2 さらに40秒間隔2回の光刺激による膜電位変化を測定した(図3)。NBS中では初回と2回の光応答はほぼ同程度であった(A)。1回目の光刺激をNBS中で行い2回目の光刺激前に細胞外からNa+を取り除いたところ、反応の減少が認められた(B,C)。この減少の程度は、細胞外Ca2+濃度を高くすると一層顕著となった(C)。すなわち視細胞の脱感作の程度は、細胞外Na+濃度を低下させた場合、もしくは細胞外Ca2+濃度を上昇させた場合に強くなることが示された。この結果は、細胞外のNa+が脱感作を抑制する方向に作用して、細胞外のCa2+が脱感作を促進する方向に作用することを示唆しており、この機序として、Na-Ca交換輸送が想定される。これらの結果は、視細胞の明順応においてNa-Ca交換輸送によるCa2+のくみ出しが重要な働きをしていることを示唆する。 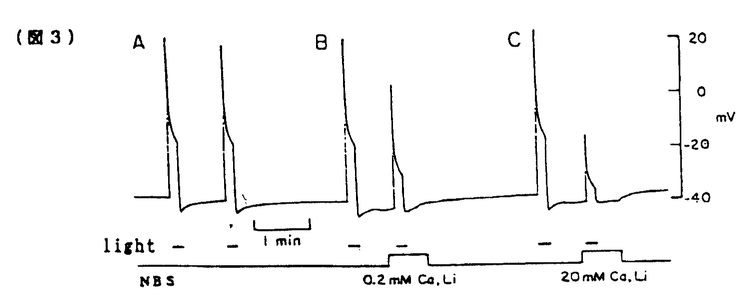 図3 図3 この点を更に明らかにする目的で、[Ca2+]1変化を測定した。 Ca2+指示薬ArsenazoIIIを細胞内に注入して、膜電位と[Ca2+]1を同時に測定した(図4)。NBS中では、光刺激による脱分極と同時に[Ca2+]1が上昇した(A)が、この上昇は0-Ca液中では、MBS中の約85.7%(n=9)であった(C)。この結果は、光刺激による[Ca2+]1の上昇には、Ca2+の細胞内storeからの放出、および細胞外からの流入の両方が関与していると考えられる。また0-Ca液中では再分極過程が緩やかであった(C)。これは、[Ca2+]1の増加の程度が、比較的小さいことに起因するかもしれない。一方、0-Na下では、暗静止状態において、[Ca2+]1の上昇が強められた(B)。これは逆向きNa-Ca交換輸送によると推定される。0-Na液中での光刺激による[Ca2+]1の上昇は、NBSとほぼ同程度であった(縦軸、対数目盛)。0-Na液での膜電位変化は、急峻な再分極、即ち強い明順応を示した。 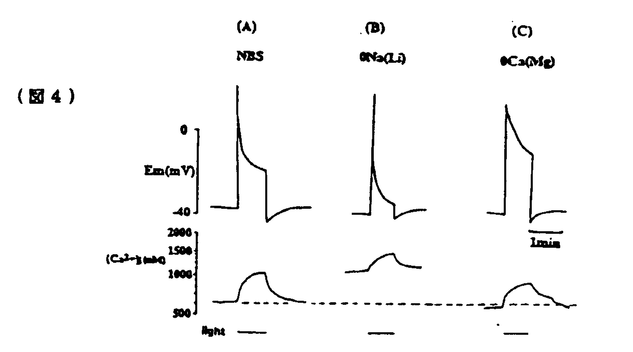 図4 図4 次にフジツボの視細胞におけるCa-H交換輪送の存在を検討した。暗状態で、細胞外液のpHを7.50から9.35にすると[Ca2+]1の上昇がみられた(図5A)。0-Na下で[Ca2+]1が上昇後一定に達した後、pHを9.35にすると、[Ca2+]1の上昇が更に認められた(B)。これに対して、0-Ca下でpHを変化させた場合には、[Ca2+]1の上昇は認められなかった(C)。この結果は、細胞外pHの上昇による[Ca2+]1の増加は、細胞外Ca2+の流入によることが示唆される。細胞外pHの上昇によって、細胞内H+が流出し、これに伴って、細胞外Ca2+がCa-H交換輸送によって流入することが示唆される。このメカニズムは、脊椎動物のrodと同様のメカニズムが、フジツボの視細胞にも存在すると考えられる。 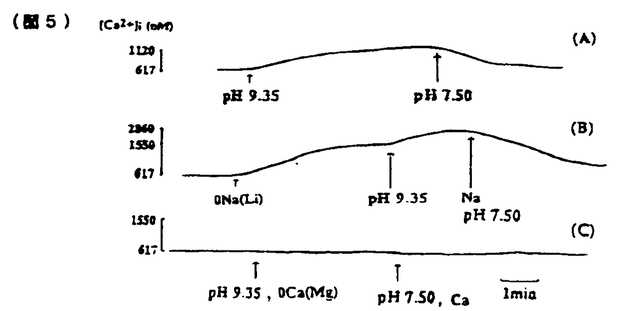 図5 図5 以上の結果より、フジツボの視細胞の光感受性は[Ca2+]1により調節されており、[Ca2+]1の恒常性は、主としてNa-Ca交換輸送により保たれていると考えられる。 またフジツボの視細胞では更に、Ca-H交換輸送が、[Ca2+]1の恒常性に寄与している可能性が示唆された。 |