非アドレナリン、非コリン作動性である知覚神経C-fiberには、substance P、neurokinin A(NKA)、neurokinin Bの三種類のタヒキニンが存在することが知られており、又、気道組織にはsubstance PやNKAを含む神経系が広く分布していることが明らかになってきている。これらのタヒキニンは、気管平滑筋の収縮の他、毛細血管からの血漿漏出、分泌線刺激など、気管支喘息にみられる臨床所見に一致する作用を示すことが報告されており、気管支喘息患者での神経性炎症のメディエイターとして働き、発作を誘発するのではないかと推察されている。また、タヒキニンは、種々の平滑筋に存在するイオンチャネルに影響を与えることが報告されているが、気管平滑筋の電気的活動に及ぼす効果については不明であった。そこで、タヒキニンの気管平滑筋の電気的活動に及ぼす効果を明らかにするため、モルモットの単離気管平滑筋細胞を用い、NKAの膜電位や膜電流に対する効果につき検討した。さらに、NKAの細胞内情報伝達機構についても検討するとともに、Ca2+依存性Cl-電流の活性化機序についても検討した。モルモット気管をコラゲナーゼ処理し、単一気管平滑筋細胞を作製、パッチ電極を用いたwhole cell clampを行った。各種薬剤は、灌流液中あるいはパッチ電極内に添加した。Tyrode液の組成(mM)は、NaCl 136.5,KCl 5.4,MgCl20.53,glucose 5.5,HEPES-NaOHbuffer 5,pH7.4であった。外液のNa+濃度を変える時には、NaClの代りに当モルのTris Cl或いはTEA Clを使用し、Cl-濃度は一定に保った。外液あるいは内液のCl-濃度を変える時には、Cl-をaspartate-で置換した。細胞膜電位やK+電流を記録する際の電極内液組成(mM)は、KCl 140,EGTA 0.15,MgCl2 2.Na2ATP 1,guanosine 5’-triphosphate(sodium salt,Sigma)0.1,HEPES-KOH buffer 5,pH 7.2であった。また内向き電流の記録のためK+電流をプロックする時には、内液のKClはCsClで置換した。さらに、電位依存性Ca2+電流の記録には、CsCl 140,MgCl2 2, HEPES-KOH 5,pH 7.2の細胞内液を用いた。 単離気管平滑筋細胞は静止膜電位が約-40mVで自発的な振動がみられたが、NKA(1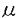 M)を投与すると膜電位は、急速に脱分極した後、-20mVまでに再分極し、その後この脱分極を維持した。保持電位-60mVにて膜電位固定しNKAを投与すると、一過性の内向きの電流を生じた。このNKA誘発電流の電流電圧曲線は、-80mVから+40mVまでの電位ではほぼ直線的な電流の増加を示し、その逆転電位は約-10mVであった。この電流は非選択性陽イオンチャネルあるいはCl-チャネルを通って主に電流が流れたものと考えられ、この内向き電流の一過性の活性化により初期の膜電位の脱分極が起こると考えられた。一方、NKA投与後の脱分極持続相でのランプパルスで見られる電流は、-40mV以上の電位において明らかに抑制された。抑制された成分をコントロールの電流トレースから差し引くことにより求めると、遅延整流特性を示し、その逆転電位は、約-70mVであった。このことより、脱分極で誘発されたK+電流がNKA投与で抑制された可能性が示唆された。実際、モルモットの気管平滑筋には、脱分極パルスで誘発されるK+電流が3種類(Ir,Is,Io)あるが、これらはNKAにより抑制を受けた。 M)を投与すると膜電位は、急速に脱分極した後、-20mVまでに再分極し、その後この脱分極を維持した。保持電位-60mVにて膜電位固定しNKAを投与すると、一過性の内向きの電流を生じた。このNKA誘発電流の電流電圧曲線は、-80mVから+40mVまでの電位ではほぼ直線的な電流の増加を示し、その逆転電位は約-10mVであった。この電流は非選択性陽イオンチャネルあるいはCl-チャネルを通って主に電流が流れたものと考えられ、この内向き電流の一過性の活性化により初期の膜電位の脱分極が起こると考えられた。一方、NKA投与後の脱分極持続相でのランプパルスで見られる電流は、-40mV以上の電位において明らかに抑制された。抑制された成分をコントロールの電流トレースから差し引くことにより求めると、遅延整流特性を示し、その逆転電位は、約-70mVであった。このことより、脱分極で誘発されたK+電流がNKA投与で抑制された可能性が示唆された。実際、モルモットの気管平滑筋には、脱分極パルスで誘発されるK+電流が3種類(Ir,Is,Io)あるが、これらはNKAにより抑制を受けた。 次に、電極内に140mMのCsClの細胞内液を充填して、NKAで一過性に活性化される内向き電流のイオン機序を調べた。電極内液のK+をCs+で置換してもこの内向き電流の活性化は見られた。この電流の逆転電位は、外液Na+濃度の変化に影響を受けなかったが、細胞内外のCl-イオンをaspartate-で置換するとCl-の平衡電位にほぼ従って変化した。さらにNKAで誘発される電流は、Cl-チャネル阻害薬である4.4’-diisothiocyanatosilbene-2,2’-disulfonic acid(1mM)で抑制された。以上より、NKAにより一過性に活性化された電流は、Cl-電流と考えられた。 そこで、NKAの膜電流に及ぼす効果に対するneurokinin受容体拮抗薬FK224の作用につき検討した。FK224(10M)自身では、コントロールの膜電流に明らかな作用はきたさないが、FK224(10M)存在下では、NKA(1M)の膜電流に及ぼす効果をほぼ完全に阻害し、Cl-電流の活性化及びK+電流の抑制効果も見られなかった。FK224を洗い流した後には、NKA(1M)投与により、Cl-電流の活性化及びその後のK+電流の抑制をきたした。又、このMKA(1M)のCl-電流の活性化及びK+電流の抑制効果は、ムスカリン受容体阻害薬のアトロピン(1M)では阻害されなかった。以上よりNKAによるこれらの膜電流に対する作用は、特異的なneurokinin受容体を介するものと考えられた。 第二部では、このNKAで誘発されるCl-電流の活性化機序につき検討した。このCl-電流が、細胞内Ca2+に影響されるかどうかを調べるため、細胞内Ca2+貯蔵部位からのCa2+を放出するcaffeine、細胞内IP3及び細胞内EGTAの効果につき検討した。保持電位を-60mVとし、caffeine(10mM)を投与するとNKAと同様の一過性の内向き電流の活性化を来し、この活性化された電流の逆転電位は、約0mVであった。また、電極内よりIP3(200M)を投与すると同様の内向き電流の活性化を生じた。このcaffeine及びIP3により活性化される電流の逆転電位は、細胞内外のCl-イオンをaspartate-で置換すると変化することより、NKA同様Cl-電流と考えられた。一方、細胞内液のEGTA濃度を0.15mMから10mMに増加させると、NKA及びcaffeineによるCl-電流の活性化は完全に阻害された。以上から、NKAによるCl-電流は、Ca2+依存性Cl-電流であると考えられ、その活性化にはIP3の関与が示唆された。実際、NKAによるCl-電流の活性化は、細胞内にIP3受容体阻害薬であるヘパリン(200 mg/ml)を充填すると完全に阻害された。次に、細胞内Ca2+貯蔵部位の関与を明らかにするため、細胞外Ca2+の影響について検討した。細胞外にCa2+非存在化でも、最初のNKAあるいはcaffeine投与によってCl-電流の活性化を認めたが、2回目の投与では活性化が見られず、これらによるCl-電流の活性化は、細胞内Ca2+貯蔵部位からのCa2+放出によると考えられた。又、細胞外にCa2+非存在化で、caffeineあるいはNKA投与によりCl-電流の活性化後、引き続きそれぞれNKAあるいはcaffeineを投与してもCl-電流の活性化はほとんど認めず、NKA及びcaffeineによるCa2+放出部位は、気管平滑筋においては、多くは一致していることが示唆された。実際、ryanodine(10 M)+caffeine処理により、その後の、caffeineやNKAによるCl-電流の活性化は著明に阻害された。さらにG蛋白の関与についても検討した。細胞内にCDP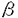 S(2mM)を投与すると、caffeineによるCl-電流の活性化は、阻害されなかったが、NKAによる活性化を完全に阻害するとともに、非水解性GTP analogueであるGTP7Sを投与するとNKAによるCl-電流の活性化を促進した。Pertussis-toxin (PTX)処理では、NKAによるCl-電流の活性化を阻害できなかった。以上より、NKAは、PTX非感受性C蛋白質の活性化によるIP3を介した細胞内Ca2+貯蔵部位からのCa2+放出によるCl-電流を活性化すると考えられた。 S(2mM)を投与すると、caffeineによるCl-電流の活性化は、阻害されなかったが、NKAによる活性化を完全に阻害するとともに、非水解性GTP analogueであるGTP7Sを投与するとNKAによるCl-電流の活性化を促進した。Pertussis-toxin (PTX)処理では、NKAによるCl-電流の活性化を阻害できなかった。以上より、NKAは、PTX非感受性C蛋白質の活性化によるIP3を介した細胞内Ca2+貯蔵部位からのCa2+放出によるCl-電流を活性化すると考えられた。 モルモットの気管平滑筋には電位依存性L型Ca2+チャネルの存在することが知られており、この活性化によりCa2+が流入してもCl-電流が活性化される可能性がある。そこで、Ca2+依存性Cl-電流の電位依存性L型Ca2+電流に伴う活性化とNKAの作用につき検討した。細胞内にEGTAを含まないCsClの細胞内液で、保持電位-60mVより+20mVへの脱分極パルスを与えると電位依存性L型Ca2+電流の活性化に伴い脱分極パルス中では、外向き電流が、-60mVの保持電位に再分極すると内向きの末尾電流が見られた。この電流は、Bay K 8644でCa2+電流を増強すると増大し、逆に外液のCa2+を除去するか、nifedipine、Cd2+によりCa2+電流をブロックすると完全に消失した。又、電極内にEGTA5 mMを充填すると消失することよりCa2+依存性の電流と考えられた。このCa2+電流により活性化される電流の逆転電位も外液のNa+を除去しても変わらず、細胞内外のCl-をaspartate-で置換するとCl-の平衡電位の変化にほぼ従って変化した。さらに、DIDS、niflumic acidは、この電流を完全に阻害したことより、これもCa2+依存性Cl-電流と考えられた。NKA及びcaffeineは、一過性にCa2+依存性Cl-電流を活性化したのち、この電位依存性L型Ca2+電流に伴うCl-電流は、むしろ著明に抑制した。この機序として、NKA及びcaffeineによる電位依存性Ca2+電流の抑制作用とともに、Ca2+貯蔵部位からのCa2+放出の関与が考えられた。即ち、心筋及び骨格筋で良く知られているCa2+-induced Ca2+release機構が、この気管平滑筋における電位依存性L型Ca2+電流によるCl-電流の活性化に関与している可能性が示唆された。また、このCa2+依存性Cl-電流は、活動電位の後電位とプラトーの形成に関与していることが確かめられた。 以上から、NKAは、モルモット単離気道平滑筋に対して急速な脱分極とその後の持続的な脱分極を来すが、これは、一過性のCa2+依存性Cl-電流の活性化とその後のK+電流の持続的な抑制によると考えられた。さらに、この脱分極によって生ずる電位依存性Ca2+電流の活性化が、NKAによる気道平滑筋の収縮反応に一部関与すると考えられた。またNKAによるCa2+依存性Cl-電流の活性化は、IP3を介した細胞内Ca2+貯蔵部位からのCa2+放出による事が明らかとなった。 |