| 内容要旨 | | 緑膿菌(Pseudomonas aeruginosa)は免疫力が低下した患者に感染症を引き起こす病原薗で,多くの抗菌薬に抵抗性を示すことから,メチシリン耐性黄色ブドウ球菌(MRSA)と並んで細菌感染症の中で最も重視されている菌種である。現在世界で使用されている抗菌薬の中で緑膿菌感染症に効果がある薬は数種類しかなく,それらはカルバペネム剤,セフェム剤,キノロン剤あるいはアミノグリコシド剤のいずれかに属している。さらにこれらの抗緑膿菌薬の臨床使用に伴い,抗緑膿菌薬に対しても耐性化した突然変異株が臨床から分離され,問題になっている。一方,in vitroでキノロン剤に耐性化した緑膿菌を選択すると,作用点や構造が異なる薬剤に対して同時に耐性化した変異株が分離されることが報告されている。このような多剤耐性緑膿菌によって感染症が引き起こされた場合は,抗菌薬の選択が難しく治療が困難である。そこで私は多剤耐性緑膿菌の耐性メカニズムを明らかにする目的で以下の検討を行なった。 多剤耐性化を引き起こすキノロン耐性遺伝子として,nalB,nfxBおよびnfxCの3種類が染色体上の独立した位置に存在することが明らかにされている。私はまず最も広範な薬剤に交差耐性を示すNalB変異株に着目し,キノロン剤であるオフロキサシンとセフェム剤であるセフスロジンを1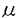 g/mlずつ含む寒天平板上で,緑膿菌標準株PAO1からNalB型の多剤耐性変異株を選択分離した。得られた変異株OCR1の薬剤感受性を表1に示す。NalB型変異株ではオフロキサシン,セフスロジンに加えカルベニシリン,テトラサイクリン,クロラムフェニコールおよびカルバペネム剤であるメロペネムに対する感受性が,親株より4から8倍低下していた。一方,メロペネム以外のカルバペネム剤(イミペネム,パニペネム)およびアミノグリコシド剤であるゲンタマイシンに対する感受性は親株とほぼ同じであった。親株および変異株から外膜画分を調製し,SDS-ポリアクリルアミドゲル電気泳動により外膜蛋白質の変化を調べたところ,NalB型変異株では見かけの分子量約50,000の外膜蛋白質(OprMと命名)の産生量が親株より増加していた。ところでnalB遺伝子とプロリン合成に関わるproC遺伝子は,染色体上の位置が近いため形質導入の際に一定の確率で同時に他の株に伝達されることが報告されている。そこでファージF116Lを用いて,分離したNalB型変異株を供与菌,PAO969(proC130)を受容菌として形質導入を行なったところ,得られたProC+形質導入株の24%がNalB変異株と同じ薬剤耐性パターンを示した(表1,T36)。したがって分離した変異株はnalB遺伝子に変異を持つ可能性が強い。また多剤耐性化とOprMの過剰産生という二つの表現型は形質導入により常に同時に伝達された。さらにOprMの過剰産生を伴う多剤耐性株が臨床分離株中に約6%存在していた。以上の結果から,OprMの過剰産生が多剤耐性化に関与していることが明らかになった。 g/mlずつ含む寒天平板上で,緑膿菌標準株PAO1からNalB型の多剤耐性変異株を選択分離した。得られた変異株OCR1の薬剤感受性を表1に示す。NalB型変異株ではオフロキサシン,セフスロジンに加えカルベニシリン,テトラサイクリン,クロラムフェニコールおよびカルバペネム剤であるメロペネムに対する感受性が,親株より4から8倍低下していた。一方,メロペネム以外のカルバペネム剤(イミペネム,パニペネム)およびアミノグリコシド剤であるゲンタマイシンに対する感受性は親株とほぼ同じであった。親株および変異株から外膜画分を調製し,SDS-ポリアクリルアミドゲル電気泳動により外膜蛋白質の変化を調べたところ,NalB型変異株では見かけの分子量約50,000の外膜蛋白質(OprMと命名)の産生量が親株より増加していた。ところでnalB遺伝子とプロリン合成に関わるproC遺伝子は,染色体上の位置が近いため形質導入の際に一定の確率で同時に他の株に伝達されることが報告されている。そこでファージF116Lを用いて,分離したNalB型変異株を供与菌,PAO969(proC130)を受容菌として形質導入を行なったところ,得られたProC+形質導入株の24%がNalB変異株と同じ薬剤耐性パターンを示した(表1,T36)。したがって分離した変異株はnalB遺伝子に変異を持つ可能性が強い。また多剤耐性化とOprMの過剰産生という二つの表現型は形質導入により常に同時に伝達された。さらにOprMの過剰産生を伴う多剤耐性株が臨床分離株中に約6%存在していた。以上の結果から,OprMの過剰産生が多剤耐性化に関与していることが明らかになった。  表1 NalB型変異株および多剤耐性形質導入株の薬剤感受性 表1 NalB型変異株および多剤耐性形質導入株の薬剤感受性 NalB変異株と同様にNfxBおよびNfxC変異株も,見かけの分子量50,000〜54,000の外膜蛋白質を親株より過剰産生していることが,異なる親株から分離した変異株で報告されている。そこでこれらの外膜蛋白質が同一の蛋白質であるか否かを明らかにする目的で,同じ親株からNalB,NfxB,およびNfxCと同じ薬剤耐性パターンを示す3種類の変異株を分離し検討を行った。PAO1からはNfxC型変異株を分離できなかったので,薬剤感受性がPAO1に類似した臨床分離株1008および8380を親株に用いた。既知の各種変異株の薬剤耐性パターンをもとにして,NalB型変異株はオフロキサシンとセフスロジンをそれぞれ1.5および3g/ml含む寒天平板上で,NfxB型変異株はオフロキサシンを1.5g/ml含む寒天平板上で,NfxC型変異株はオフロキサシンとイミペネムを1.5g/mlずつ含む寒天平板上で,それぞれ選択分離した。NfxB型変異株はキノロン剤に加えセフピロムやセフォゾプランなどの新しい双性イオン性のセフェム剤に交差耐性を示したが,他の -ラクタム剤には交差耐性を示さなかった。NfxC型変異株はカルバペネム剤とキノロン剤に交差耐性を示した。3種類の多剤耐性変異株の外膜蛋白質を比較すると,NalB型変異株では見かけの分子量約50,000の外膜蛋白質(OprM)の産生量が,NfxB型変異株では見かけの分子量約54,000の外膜蛋白質の産生量が,それぞれ親株より増加していた。またNfxC型変異株ではカルバペネム剤の選択的透過孔を形成する見かけの分子量約46,000の外膜蛋白質OprDの産生量減少と見かけの分子量がOprMと同じ外膜蛋白質の産生量増加が見られた。しかし泳動前の蛋白質の可溶化を37℃で行うと,NalB型変異株で産生量が増加したOprMの泳動度は見かけの分子量100,000以上の高分子量側に変化したが,NfxC型変異株で産生量が増加した外膜蛋白質の泳動度は変化しなかった。したがって両者は異なる蛋白質であると考えられる。そこでNfxBおよびNfxC型変異株で産生量が増加した蛋白質をそれぞれOprJおよびOprNと命名した。以上の結果から,少なくとも3種類の異なる外膜蛋白質OprM,OprJおよびOprNが多剤耐性化に関与していることが明らかになった。 -ラクタム剤には交差耐性を示さなかった。NfxC型変異株はカルバペネム剤とキノロン剤に交差耐性を示した。3種類の多剤耐性変異株の外膜蛋白質を比較すると,NalB型変異株では見かけの分子量約50,000の外膜蛋白質(OprM)の産生量が,NfxB型変異株では見かけの分子量約54,000の外膜蛋白質の産生量が,それぞれ親株より増加していた。またNfxC型変異株ではカルバペネム剤の選択的透過孔を形成する見かけの分子量約46,000の外膜蛋白質OprDの産生量減少と見かけの分子量がOprMと同じ外膜蛋白質の産生量増加が見られた。しかし泳動前の蛋白質の可溶化を37℃で行うと,NalB型変異株で産生量が増加したOprMの泳動度は見かけの分子量100,000以上の高分子量側に変化したが,NfxC型変異株で産生量が増加した外膜蛋白質の泳動度は変化しなかった。したがって両者は異なる蛋白質であると考えられる。そこでNfxBおよびNfxC型変異株で産生量が増加した蛋白質をそれぞれOprJおよびOprNと命名した。以上の結果から,少なくとも3種類の異なる外膜蛋白質OprM,OprJおよびOprNが多剤耐性化に関与していることが明らかになった。 NfxB変異株の薬剤感受性パターンは株によって若干の相違があった。この相違の理由を明らかにする目的で,PAO1から複数のNfxB型変異株を分離し薬剤感受性を詳細に検討した。NfxB型変異株はセフピロムとオフロキサシンをそれぞれ2および1g/ml含む寒天平板上で選択分離した。分離したNfxB型変異株を感受性変化の程度に基づいてタイプAとタイプBに分類した。タイプA変異株COR1は,セフピロムおよびオフロキサシンに対して親株より8倍耐性化していた(表2)。一方,タイプB変異株COR4はセフピロムおよびオフロキサシンに加えてテトラサイクリンおよびクロラムフェニコールに対しても親株より耐性化しており,逆にカルベニシリン,イミペネムおよびゲンタマイシンに対し親株より4倍高感受性化していた。全体を通じてタイプB変異株はタイプA変異株より薬剤感受性変化の程度が大きかった。野生型nfxB遺伝子を含むプラスミドpNF111をNfxB変異株に形質転換すると,nfxB遺伝子の変異が相補され親株と同じ薬剤感受性を示すことが報告されている。タイプAおよびタイプB変異株の薬剤感受性はpNF111で形質転換することにより,親株の感受性レベルに回復した。したがってこれらの変異株はいずれもNfxB変異株であることが確認された。両タイプのNfxB変異株から外膜蛋白質を調製し,OprJに対するモノクローナル抗体を用いてイムノブロット解析を行ったところ,タイプB NfxB変異株はタイプA NfxB変異株よりもOprJ産生量が多く,親株PAO1のOprJ産生量は検出限界以下であった。さらに染色体伝達能をもつプラスミドFP5を用いて,各タイプのNfxB変異株を供与菌として接合を行い,変異nfxB遺伝子が伝達された接合体を分離した。得られた接合体はそれぞれの対応する供与菌とほぼ同じ薬剤感受性パターンおよびOprJ産生量を示した。以上の結果から,異なる表現型を示す複数のnfxB変異が存在することが明らかになった。またNfxB変異株における薬剤感受性変化の程度とOprJ産生量が相関していることが明らかになった。  表2 NfxB変異株の薬剤感受性 表2 NfxB変異株の薬剤感受性 |