ホメオスタシスの維持と自律機能の制御は個体の維持のみならず種の保存に必要不可欠な要素であり、少なくとも哺乳類において視床下部はその最高中枢であることが知られている。視床下部は内分泌、自律神経、情動などを介した遠心性制御と、フィードバックを中心とする求心性の情報を統合し、生体内環境の動的平衡を常に保ち続けていると考えられている。しかし、この動的平衡の設定値は常に一定にあるわけではなく、生体内環境を長期的な時間軸に展開してみると日周性、成長、老化など、個体が免れることができない普遍的な法則に基づいて変化している。今日まで、視床下部の生理機能の探求を目的として、薬物投与や電気刺激などによる急性的な反応を指標にして主に研究が進んできたが、この様な普遍的な中長期的変化とその調節機構を理解してゆくこともまた生理学の重要な目的の一つであるといえる。本論文では思春期発来と概日リズムの発現に焦点を当て、視床下部レベルでの長期的な調節機序について検討を試みた。 霊長類における性成熟の成立には、視床下部からの黄体形成ホルモン放出ホルモン(LHRH)の分泌上昇が必要不可欠であり、性成熟の開始前にLHRHニューロンが既に形態的にも機能的にも完成されていることから、LHRHニューロンの脱抑制もしくは興奮性制御の成立が性成熟の起因であるとの仮説が示されてきた。本論文第1章では性成熟過程での雌アカゲザルのLHRH分泌に対するGABAニューロンの役割について、覚醒無麻酔条件下でのpush-pull perfusion法により検討した。実験には前性成熟期または性成熟期の雌アカゲザルを用い、正中隆起部に脳定位的にpush-pull cannulaを植え込んだ。実験当日に正中隆起部を人工脳脊髄液で局所的に灌流しつつ、フラクションコレクターを用いて10分毎にサンプリングした。灌流液中のLHRH濃度はラジオイムノアッセイ(RIA)によって測定し、GABA濃度は高速液体クロマトグラフィー(HPLC)によって測定した。結果として、1)正中隆起へのGABAの投与(10-5M及び10-7M)がLHRH分泌を抑制する。2)前性成熟期では性成熟期と比べ正中隆起でのGABAの分泌レベルが高い。3)GABAA受容体のアンタゴニストであるビキュキュリン(10-5M及び10-7M)は特に前性成熟期においてLHRH分泌を強く刺激する一方、GABAB受容体のアンタゴニストであるハイドロキシサクロフェン(10-5M及び10-7M)では有意な効果が認められないことが明らかとなった。以上より、前性成熟期には正中隆起から分泌されるGABAがGABAA受容体を介してLHRHの分泌上昇とそれに続く性成熟の開始を強く抑制していることが示唆された。 GABAは2種類の酵素、GAD67とGAD65によってグルタミン酸を基質として生産されることが知られているが、ヒトにおけるこれらの酵素の全塩基配列が近年決定された。そこで第2章ではまずアカゲザルの視床下部3領域(視索前野、内側底部視床下部、後視床下野)における各GADmRNAの存在をヒトでの塩基配列を基にPCR法を使って確認し、両GADmRNAともに視床下部の3領域に存在することが判明した。次に、前性成熟期のアカゲザルの内因性GABAの生産を阻害する目的で、各GADmRNAの開始コドン領域に相補的な20mer-アンチセンスオリゴヌクレオチドを合成して人工脳脊髄液で溶解し(10-6M)、これをpush-pull perfusion法を用いて正中隆起に6時間連続的に灌流投与した。コントロール群には塩基配列を入れ替えた20merスクランブルドオリゴヌクレオチドを使用した。灌流液中のLHRH濃度をRIAによって測定したところ、両GADmRNAに対するアンチセンスはともに有意にLHRHの分泌を刺激するが、スクランブルドオリゴでは効果が認められないこと、CAD67mRNAに対するアンチセンスはGAD65mRNAよりも効果が強いことが明らかとなった。以上より、霊長類の性成熟過程におけるGADmRNAの発現もしくはGADの活性変化が性成熟開始の要因であることが示唆された。 LHRHの分泌上昇はGABAニューロンからの脱抑制のみでなく、何らかの興奮性入力の上昇によってもたらされた可能性も考えられる。特に成熟アカゲザルにおけるニューロペプチドY(NPY)のパルス状分泌はパルス状LHRH分泌と強く相関していること、抗NPY抗体を正中隆起に直接投与するとLHRHのパルス状分泌が抑制されることが報告されている。第3章では、性成熟過程でのLHRH分泌におけるNPYニューロンの役割を同様にpush-pull perfusion法を用いて検討した。灌流液中のNPY濃度はRIAによって測定した。結果として、正中隆起におけるパルス状のNPY分泌は前性成熟期では比較的低いが性成熟期では高まること、正中隆起への抗NPY抗体(100倍及び1000倍希釈)の直接投与は、前性成熟期ではLHRH分泌に対して効果が認められないが、性成熟期では有意にLHRH分泌を抑制することが判った。従って、GABAとは逆に、内因性のNPYによるLHRHニューロンに対する興奮性の入力は性成熟期以降に上昇し、性成熟期以降のパルス状LHRH分泌の形成に貢献していることが示唆された。 神経回路網の変形法は長期的な神経機能の解析法として特に有効な手段であると考え、第4章では、この方法論をラットを用いて確立すると共に、長期的な摂食抑制機能における視床下部腹内側核(VMH)の役割について検討した。実験にはウィスター系雌性ラットを用い、生理食塩水内で飽和膨潤させた1.6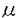 lの吸水性ハイドロポリマーゲル(PG)を脳定位的にVMH内に注入して局所的にVMHの神経回路網の変形を促した。その結果、1)両側性および片側性のVMHへのPG注入は1週間以上にわたって有意に摂食量と体重の減少を誘起する。2)微小ナイフによるVMHの全周囲切断はPG注入による摂食抑制効果を阻害する。3)GABA(160 nmol及び80 nmol)をPGと共にVMHへ注入すると濃度依存性にPGによる摂食抑制効果が減弱される。4)片側性にPGをVMH内に注入すると、PG注入側のVMHニューロンの自発発火頻度は対側のVMHニューロンよりも有意に自発発火頻度が上昇する。5)片側性にPGをVMH内に注入すると、特にPG注入側のVMHニューロンで転写調節因子の一つであるFos蛋白の発現が認められることが明らかとなった。以上より、PGのVMH内への注入はVMH内ニューロンの長期的神経出力を誘発すること、また、VMH内での長期的な摂食抑制作用を持つニューロン群の存在が示唆された。 lの吸水性ハイドロポリマーゲル(PG)を脳定位的にVMH内に注入して局所的にVMHの神経回路網の変形を促した。その結果、1)両側性および片側性のVMHへのPG注入は1週間以上にわたって有意に摂食量と体重の減少を誘起する。2)微小ナイフによるVMHの全周囲切断はPG注入による摂食抑制効果を阻害する。3)GABA(160 nmol及び80 nmol)をPGと共にVMHへ注入すると濃度依存性にPGによる摂食抑制効果が減弱される。4)片側性にPGをVMH内に注入すると、PG注入側のVMHニューロンの自発発火頻度は対側のVMHニューロンよりも有意に自発発火頻度が上昇する。5)片側性にPGをVMH内に注入すると、特にPG注入側のVMHニューロンで転写調節因子の一つであるFos蛋白の発現が認められることが明らかとなった。以上より、PGのVMH内への注入はVMH内ニューロンの長期的神経出力を誘発すること、また、VMH内での長期的な摂食抑制作用を持つニューロン群の存在が示唆された。 当教室ではこれまで、VMHが行動賦活系の一つであることを示してきた。近年、行動の発現自身が長期的な行動概日リズムを調節することが報告され、このことから脳内の行動制御中枢による概日リズムの発現制御の可能性が考えられる。第5章では行動概日リズム発現が、行動賦活系の一つであるVMHによって調節されている可能性を考え、第4章で確立したVMH内神経回路網の変形による持続的な神経出力誘発法を用いて検討した。その結果、1)VMHへのPG注入(1.6l)は行動概日リズムを著しく減弱させるが、視索前野や後視床下野へのPG注入では明確な行動概日リズムの発現が認められる。2)VMHへのPG注入は正常性周期に対してはこれを停止して連続発情状態を促し、血中メラトニンの日内変動に対しては主観的暗期における分泌上昇を抑制して有意な日向変動を消失させる。3)微小ナイフによるVMHの前方切断および後方切断はVMHへのPG注入効果を阻止しないが、VMHの全周囲切断および上方切断はVMHへのPG注入効果を阻止して行動概日リズムの発現を促すことが明らかとなった。従って、VMH内には概日リズム発現を調節するニューロン群が存在し、この神経出力はVMHから上方性に投射していることが示唆された。 本論文では視床下部における持続的な神経性調節機能についてpush-pull perfusion法と神経回路網の変形法を用いて検討を試みた。今日まで、霊長類の性成熟の発来に必要不可欠の要素である正中隆起におけるLHRH分泌の上昇がどの様な神経性入力の変化によって誘起されるのか明らかではなかったが、第1章から第3章の実験でGABAA受容体を介したGABAニューロンからの脱抑制とNPYニューロンによる興奮性入力の増大がLHRH分泌の上昇とそれに続く春期発来を誘起することが示唆された。また、第4章と第5章では、PGの微量注入による神経回路綱の変形法を確立し、PGのVMH内注入がVMH内ニューロンからの持続的な神経出力を誘発することが明らかとなった。この方法を用いて、VMH内での長期的に摂食行動を抑制するニューロン群と概日リズム発現を抑制するニューロン群の存在が示唆された。これまでに給餌条件や老化によって行動概日リズムの発現が弱まることが報告されているが、これらのニューロン群が担う生理的意義については更なる検討が必要である。 |