本研究は、乳癌細胞におけるヒトc-erbB-2遺伝子の過剰発現のメカニズムおよび、c-erbB-2遺伝子プロモーターに結合する因子としてクローニングしたFnb蛋白質についての解析を試みたものであり、下記の結果を得ている。 1.乳癌細胞においてヒトc-erbB-2遺伝子が過剰発現している原因の少なくとも1つに、c-erbB-2遺伝子の転写速度の上昇があることを明らかにした。 2.乳癌細胞を用いてヒトc-erbB-2遺伝子のプロモーター解析を行い、ヒトc-erbB-2遺伝子プロモーターが複数の転写因子による制御を受けていることを示した。この転写因子には、細胞株特異的なものや非特異的なもの、活性化因子や抑制因子、CCAAT box結合蛋白質が含まれることが明らかになった。 3.ヒトc-erbB-2遺伝子高発現乳癌細胞株MDA-MB453細胞とHeLa細胞から調製した核抽出液を用いてゲルシフトアッセイを行ったところ、c-erbB-2遺伝子プロモーター断片には、種々の核蛋白質が結合することが明らかになり、プロモーター解析から示唆された蛋白質が存在する可能性を示した。 4.ヒトc-erbB-2遺伝子プロモーター断片への結合活性を指標としてfnb遺伝子をクローニングした。fnb遺伝子は、全長5.8kb、1656アミノ酸のフィンガー蛋白質をコードしていた。Fnb蛋白質は分子量250kDであり、計算値184kDとの差はカルボキシル末端領域の寄与が大きいことが示された。さらに、その結合領域はc-erbB-2遺伝子プロモータ-+5から+36位の領域(K領域)であることが示された。 5.fnb遺伝子は、調べた全ての細胞株で弱く発現しており、c-erbB-2遺伝子過剰発現細胞株と他の細胞株との違いは見出されず、また、血球系の細胞では発現しないc-erbB-2遺伝子とは発現パターンが異なっていた。 6.fnb遺伝子発現ベクターを用いた共トランスフェクションの実験により、c-erbB-2遺伝子プロモーター配列からK領域を欠失しても、ヘテロなプロモーターにK領域を挿入しても、Fnb蛋白質による転写活性化には影響がないことが示された。従ってFnb蛋白質が、直接単独でc-erbB-2遺伝子プロモーターに作用している可能性は低いと考えられるが、一方、ヒトメタロチオネインIIAプロモーター、単純性疱疹ウイルスチミジンキナーゼプロモーターからの転写を活性化し、転写活性化因子としての機能を有することが明らかにされた。 7.全長のFnb蛋白質は核内のnuclear bodiesに局在するが、アミノ末端側を欠失することにより、その局在は損なわれることが示された。 以上、本論文は乳癌細胞におけるヒトc-erbB-2遺伝子の過剰発現のメカニズムを検討し、新規の転写因子Fnb蛋白質をクローニングし、機能解析を行った。本研究はこれまで解析の遅れていた乳癌細胞におけるヒトc-erbB-2遺伝子の転写制御に関する研究を集中的に行い、さらに新規の遺伝子をクローニングしており、学位の授与に値するものと考えられる。 | 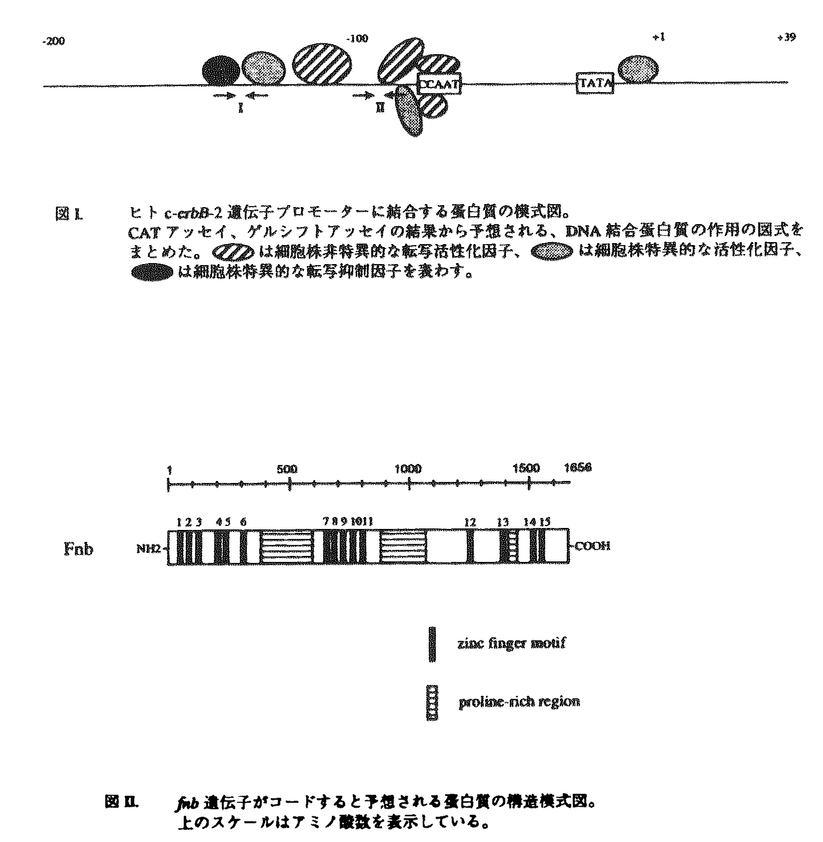
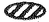 は細胞株非特異的な転写活性化因子、
は細胞株非特異的な転写活性化因子、 は細胞株特異的な活性化因子、
は細胞株特異的な活性化因子、 は細胞株特異的な転写抑制因子を表わす。 / 図II. fnb遺伝子がコードすると予想される蛋白質の構造模式図。 上のスケールはアミノ酸数を表示している。
は細胞株特異的な転写抑制因子を表わす。 / 図II. fnb遺伝子がコードすると予想される蛋白質の構造模式図。 上のスケールはアミノ酸数を表示している。