| 内容要旨 | | 棘皮動物は水管系と呼ばれる独特の器官系を持つ。棘皮動物の管足は水管系の一器官で、移動・摂食・呼吸などの生活上の重要なはたらきをになっている。水管系の内腔は体腔のひとつであるが、主体腔が閉じた系であるのに対して外の海水につながっている。この水管系と外界をつなぐ器官が多孔体(madreporite)である。多孔体は骨格と軟組織からなり、多孔体の多数の骨の孔には通水管(pore canal)と呼ばれる内面が繊毛上皮でおおわれている管がはまりこんでいる。多孔体は棘皮動物の水管系が機能するために何らかの重要な働きをしていると考えられており、19世紀以来多くの研究者が多孔体についてさまざまな観察と実験を行ってきた。それにもかかわらず、多孔体の機能については一致した見解が得られていない。その原因は主に、多孔体を通る水流の方向について一致した結果が得られていないことによる。何人かの研究者は多孔体を通って水管系内に海水が入ると主張している(Ludwig,1890;MacBride,1896;Gemmill,1914;Budington,1942;Bargmann and Behrens,1964;Ferguson,1984,1988,1989,1990,1992)。一方、別の研究者は多孔体を通る水流の方向は外向きで、多孔体は排出器官として働くと主張している(Hartog,1887;Ruppert and Balser,1986)。またある者は多孔体は水流をおこすためにあるのではなく、外界と水管系内との圧力を等しく保つ器官であると言っている(Fechter,1965)。多孔体の機能を明らかにすることは棘皮動物という動物群を理解する上で重要であると考え、ウニを材料として本研究を行った。 本研究は主に1.単離した多孔体を用いた実験2.単離した通水管を用いた実験3.透過型電子顕微鏡による観察4.ローダミン・ファロイジン染色の4つの部分からなる。単離した多孔体を用いた実験では生きた状態で単離した多孔体を上から観察して、孔の大きさが刺激に反応して変わるかどうか調べた。その結果、多孔体の孔が刺激に反応して開閉するという全く新しい知見を得た。孔の開閉の機構を知るために、単一の通水管を多孔体の孔から単離する新たな手法を開発し、単離した通水管を用いた実験を行った。また、多孔体の機能を知るためには、通水管を通る水流が有るかどうか、有るとすればそれはどちら向きかを知ることが重要であると考え、単離した通水管を用いた実験では通水管を通る水流の方向についても調べた。多孔体については、さまざまな研究がなされてきたにもかかわらず、FergusonとWalker(1991)の研究を除いて、これまで透過型電子顕微鏡を用いた研究は行われていなかった。多孔体の孔の開閉の機構を調べるためだけではなく、多孔体の機能を知るためにも、その微細構造を調べる必要があると考え、透過型電子顕微鏡による観察を行った。透過型電子顕微鏡を用いたウニの多孔体の観察は、本研究が最初の報告である。電子顕微鏡による観察の結果、通水管の細胞内に孔の開閉に関与すると思われる繊維構造が見られたので、ローダミン・ファロイジン染色を行いこの繊維がF-アクチンからなるかどうか調べた。各々の結果は以下の通りである。なお、本研究では主に三崎臨海実験所付近で採集したバフンウニ(Hemicentrotus pulcherrimus)を用いた。 1.単離した多孔体を用いた実験 多孔体をウニから切り出し、その表面を薄切した。薄切した組織を海水で満たしたトロフに表面を上にして固定し、20倍の水浸系対物レンズを用いて透過光で孔の開口部を観察した。 多孔体の孔の大きさに対する薬物の影響を調べた結果、多孔体の孔が10-8M以上の濃度のアセチルコリンの存在下で可逆的に閉じることがわかった。アセチルコリン投与前平均約20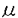 mあった孔の直径は、10-6-10-5Mのアセチルコリンを投与すると2分間で最初の大きさの平均約70%になり、6分間で平均約60%になった。 mあった孔の直径は、10-6-10-5Mのアセチルコリンを投与すると2分間で最初の大きさの平均約70%になり、6分間で平均約60%になった。 アセチルコリンの孔を閉じさせる作用は濃度依存性があり、10-6M以上の濃度で最大の反応が見られた。アセチルコリンの拮抗薬の効果を調べたところ、アセチルコリンの作用はアトロピンで可逆的に阻害されたが、d-ツボクラリンでは阻害を受けなかった。また、10-5Mのアドレナリンは孔の大きさを変化させなかった。これらの結果は多孔体の孔を囲む細胞がムスカリン様のコリン作動性支配を受けていることを示唆している。 2.単離した通水管を用いた実験(a)方法 単一の通水管は以下の方法で単離した。まず、ウニから多孔体を切り出し、その表面を薄切した。次に、多孔体の骨格と通水管の間にある基底膜を溶かすために、0.2%のコラゲナーゼで30℃で1時間組織を処理した。その後、まつ毛で作った毛針を用いて軟組織を骨から分離した。分離した軟組織は、多孔体の外表面をおおう上皮とそれにつながる通水管からなる。個々の通水管は、ガラス針を用いて外表面の上皮から切り離すことによって単離した。単離した管は微分干渉顕微鏡を用いて側面から観察した。 (b)アセチルコリンの作用による通水管の形態変化 通水管のどのような形態変化によって孔が閉じるのかを知るために、アセチルコリンの作用によって管のどの部分にどのような変化がおこるかを調べた。アセチルコリンを作用させると、管の内径と外径が小さくなり、管壁は厚くなり、管は短くなった。この反応は可逆的で、アセチルコリンを除去すると逆の形態変化がおこった。 多孔体の孔が閉じる機構のひとつとして、通水管の細胞が吸水等によって膨張して孔が閉じる可能性が考えられたので、管の各部分の計測値から孔の開閉に伴って軟組織の部分の体積が変化するかどうかを調べた。計算の結果、孔の開閉は管の軟組織の部分の体積変化を伴わないことが示唆された。この結果は、通水管の細胞が吸水等により膨張することによって多孔体の孔が閉じるのではなく、何らかの収縮構造の関与によって孔が閉じることを示唆した。 (c)通水管を通る水流に関する研究 通水管を通る水流の方向については、通水管の反口側(からだの外にあたる側)の端、もしくは口側(からだの内にあたる側)の端に墨汁を懸濁した海水をマイクロピペットを使って吹き出すことによって調べた。通水管の反口側の端に墨汁を吹き出した場合、15例中11例、墨汁は通水管の中を通って反対側から出てきた。管の口側の端に墨汁を吹き出した場合は、墨汁は通水管の繊毛にはじかれて、管の中へは入っていかなかった。墨汁の動きとは対照的に、径の大きな(約1m以上)粒子は管の中心を通って内から外へ(口側から反口側へ)運ばれることがわかった。 一方、アセチルコリンの作用で内径が小さくなっている通水管では、大きな粒子が内から外へと運ばれるのは見られたが、管のどちら側の端に墨汁を吹き出しても、墨汁が管を通って反対側から出てくることはなかった。 これらの結果は、通水管では内向きの水流があることと、その水流は管の内径が小さくなると妨げられることを示している。 3.透過型電子顕微鏡による観察 多孔体の微細構造、特に孔の開閉に関与する構造が何なのかを調べるために、透過型電子顕微鏡を用いて通水管の切片を観察した。通水管は繊毛上皮細胞で裏うちされており、管には筋肉細胞は見られなかった。管を裏うちする上皮細胞は頂端部で接着帯とseptate junctionにより隣の細胞と結合していて、細胞内には接着帯をつなぐ繊維構造が見られた。 4.ローダミン・ファロイジン染色 電子顕微鏡で通水管の細胞内に見られた繊維構造がF-アクチンからなるかどうかを調べるために、通水管の切片をローダミン・ファロイジンで染色して落射型蛍光顕微鏡で観察した。その結果、電子顕微鏡で繊維が見られた部分がローダミン・ファロイジンで強く標識されることがわかった。このことは繊維構造がF-アクチンからなることを示唆する。 以上、本研究の結果、多孔体の孔が刺激に反応して開閉するということが新たにわかった。単離した通水管を用いた実験は、孔の開閉が細胞の体積変化を伴わないことを示した。透過型電子顕微鏡による観察とローダミン・ファロイジン染色の結果は、孔が閉じる反応は、通水管を裏うちする上皮細胞の頂端部に存在するF-アクチンからなる繊維が収縮して、細胞の丈が高くなることによっておこることを示唆する。また、本研究によって、単離した通水管では反口側から口側へ向かう内向きの水流がある一方、大きな粒子はその水流とは逆方向へ運ばれることがわかった。多孔体を通る水流について、これまでの研究で一致した結論が得られなかったのは、通水管が、本研究で示したような、両方向性の物質輸送を行っているためだと思われる。通水管の繊毛がおこす水流は水管系内の液体の体積あるいは圧力を調節している可能性がある。アセチルコリンの作用で内径が小さくなった通水管では水流が見られなかったことから、多孔体を通る物質の移動は孔の開閉によって制御されていることを示唆する。これらの新しい知見によって、多孔体の構造と機能に関する多くの疑問が解明された。また、本研究で得られた結果は棘皮動物という重要な動物門の理解に貢献するものである。 |
| 審査要旨 | | 棘皮動物は水管系と呼ばれる独特の器官を持ち,この水管系が棘皮動物の生活上重要な,移動・摂餌・呼吸などの働きをになっている.多孔体は,この水管系と外界とをつなぐ器官であり,水管系にとって重要な働きをしていると考えられるがその機能は明かではない.本論文は,ウニ多孔体の機能について,単離した多孔体を用いてその生理的性質を微細構造との関連において明かにするという独創的な方法により研究したものである. 多孔体に関する研究は19世紀以来数多く行われてきたが,その機能は不明である.多孔体は水管系の入り口(あるいは出口)にあたる器官であることから,水の出入りがあるであろうと推測されている.ところが,これまでの研究では水流の方向について,外向きであるという報告と内向きであるという報告があり,一致した見解はいられていない.多孔体の機能の解明には,まずこの多孔体を通る水流の方向を明かにすることが重要である. 本研究では,主にバフンウニを用いて,1.単離した多孔体を用いた実験,2.単離した通水管を用いた実験,3.透過型電子顕微鏡による多孔体の微細構造の観察,4.通水管細胞のローダミンファロイジン染色,の4つの手法による実験を行った. 単離した多孔体を用いた実験では,多孔体をウニから切り出し,その表面を薄切して海水中で観察した.低真空透過型電子顕微鏡を用いた予備観察から,多孔体の孔の大きさが変化する可能性が示唆された.そこで,孔の大きさに対する薬物の効果を調べた結果,孔は10-8M以上のアセチルコリンの存在下で可逆的に閉じることがわかった.閉じ方は,アセチルコリンの濃度に依存していた. 孔の開閉の機構の解析と,多孔体を通る水流の方向の解析を目指して,単一の通水管を多孔体の孔から単離する新たな手法を開発した.通水管の単離には,まず多孔体を0.2%コラゲナーゼで1時間処理後まつ毛で作った毛針を用いて軟組織を骨から分離した.さらに分離した軟組織から多孔体の外表面を覆う上皮をガラス針で切り離して通水管を得た.この通水管を側面から微分干渉顕微鏡により観察した.通水管細胞が吸水などによって膨張することで多孔体の孔が閉じる可能性も考えられた.そこでアセチルコリン作用時の孔の開閉に伴う単離した通水管全体の形態変化を解析し,孔の開閉の際の軟組織の体積変化を検討した.この結果,通水管の細胞が吸水などにより膨張することによって多孔体の孔が閉じるのではないことが明かとなった.従って,何らかの収縮機構の関与が示唆された.さらに,単離した通水管を用いて通水管を通る水流の方向について検討した.墨汁を懸濁した海水を単離した通水管近くに与え墨汁の動きを解析したところ,墨汁は,反口側(多孔体の外側)から口側(内側)へと移動するが,逆方向へは移動しないこと,この移動は,アセチルコリンを作用させて孔が閉じたときには起こらないこと,一方,径が約1m以上の大きな粒子は,口側から反口側へと移動することがわかった. 孔の開閉の構造的基礎を明かにするために,通水管の切片を透過型電子顕微鏡で観察した.通水管は繊毛上皮細胞で裏打ちされており,管には筋肉細胞は見られなかった.しかし,繊毛上皮細胞は,その頂端部で接着帯とseptate junctionにより隣の細胞と結合しており,結合部分付近に繊維構造が見られた.次に,この繊維構造がF-アクチンからなるかどうかを調べる目的で,通水管の切片をローダミン・ファロイジンで染色して落射型蛍光顕微鏡で観察した.その結果,この繊維構造の部分が強く標識されたことから,電子顕微鏡で見られた繊維構造はF-アクチンからなることが示唆された. 本研究により,多孔体を通る水流について両方向性の物質輸送があることが明かとなった.また,多孔体の孔が刺激に反応して開閉することが初めて発見され,その孔の開閉が通水管を裏打ちする上皮細胞内のF-アクチンからなる繊維構造の収縮により引き起こされる可能性が示された.これらの結果は,多孔体を通る物質の移動は孔の開閉によって制御されていることを示唆する.また,通水管の繊毛が起こす水流は水管系内の液体の体積あるいは圧力を調節している可能性がある. 以上のように,本研究は,多孔体の機能について独創的な実験方法により解析し,いくつかの重要な知見を得たものであり,動物生理学の基礎的研究にとって意義があるばかりでなく,棘皮動物という重要な動物門の理解に貢献するものである.よって,本論文は博士(理学)の学位論文にふさわしいと,審査委員会委員は全員一致で認めた.なお,本論文の内容は,共同研究によるものであるが,論文提出者が主体となって実験及び解析を行い,本論文に対し論文提出者の寄与が充分であると判断された. |