本研究は、血管の発生や血管病変において重要な役割を果たしている平滑筋の役割を明らかにするために、平滑筋特異的に発現する平滑筋ミオシン重鎖(SM1/2)遺伝子について、5’プロモーター領域におけるin vitroでの発現調節機序の解析を試みたものであり、下記の結果を得ている。 1.マウス平滑筋ミオシン重鎖遺伝子の5’プロモーター領域をクローニングした。プライマー伸長法により大動脈、腸管、子宮における転写開始点は、ATG開始コドンから106bp上流の一カ所に同定された。転写開始点から5’側に1526bpの領域をシークエンスした結果、-28bpにTATA box、-43bpにGATA box、-965bpと-1297bpにCArG box、そして8個のE-boxを認めた。 2.5’プロモーター領域を種々の長さに欠失させたSM1/2プロモーターを組み込んだルシフェラーゼ・レポーター遺伝子を各種細胞に一過性に導入し、転写活性を比較した。1226bpのプロモーター領域を含むレポーター遺伝子はウサギ大動脈平滑筋細胞の初代培養細胞に導入されると、他の細胞に導入された場合より高い転写活性を示した。この高い転写活性の原因は主に、-92bpと-80bpの間と、-72bpより近位のプロモーター領域の活性増加作用によると考えられた。 3.平滑筋培養細胞の核タンパク質を用いたゲルシフト分析により、SM1/2プロモーター領域の-89bpと-69bpに存在するCCTCCC配列に特異的に結合する核タンパク質の存在が示された。このCCTCCC配列に変異を導入したレポーター遺伝子の転写活性が変異を導入されていないレポーター遺伝子より低値を示したことは、この配列がSM1/2遺伝子の発現に重要であることを証明している。 4.既知のシス・エレメントであるCACC-binding boxとSp1-binding boxを含むオリゴヌクレオチドを用いたゲルシフト分析により、CACCC結合タンパク質とSp1が、CCTCCC配列に結合できることを示した。また、Sp1に対するポリクローナル抗体を用いたゲルシフト分析で、CCTCCC配列に結合した平滑筋培養細胞の核タンパク質中にSp1が存在することが明らかになった。これまでに、CACCC結合タンパク質とSp1は、いずれも組織特異的遺伝子発現に関与している例が知られている。 5.最近報告されたウサギのSM1/2遺伝子のプロモーター領域とマウスのプロモーター領域のシークエンスを比較した。TATA boxと二連のCCTCCC配列を含む転写開始点から-120bpまでの近位プロモーター領域と、CArG boxを含む-1000bpと-1500bpの間のプロモーター領域で、類似した配列を認めた。CCTCCC配列がマウスとウサギで種を越えて保存されていることは、この配列がSM1/2遺伝子発現に重要であることを支持している。 以上、本論文は、マウス平滑筋ミオシン重鎖遺伝子の5’プロモーター領域に存在する二連のCCTCCC配列とその結合タンパク質が平滑筋特異的な遺伝子発現を規定していることを明らかにした。結合タンパク質としては、Sp1またはZinc Fingerを含む転写因子が関与している可能性が示唆される。本研究は、血管の発生や血管病変における平滑筋の役割の解明に大きく貢献するものであり、動脈硬化や血管形成術後の再狭窄病変形成の機序の解析とその治療法の開発にも役立つと期待され、学位の授与に値するものと考えられる。 | 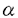 アクチン、SM22
アクチン、SM22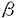 ガラクトシダーゼ・レポーター遺伝子を細胞に導入し、転写効率を補正した。まず、ウサギ大動脈平滑筋由来の血管平滑筋細胞C2/2にSM1/2が発現していることをRNase Protection Assayにて確認した後、転写活性を比較検討した。3.4kbp、1226bpのプロモーター領域を含むプラスミドの転写活性は、プロモーター・エンハンサーを含まないレポーター遺伝子、PGVB、に比較して50倍近い活性の上昇を認めた。-188bpまで欠失させたコンストラクトでも転写活性はそれほど変化しなかったが、より近位、特に、-92bpと-80bpの欠失で転写活性は顕著に低下し、この領域に活性増加作用領域が存在する可能性が示された。-72bpまで欠失させたプラスミドの転写活性はPGVBの9倍程度あるので、より近位に必須のシスエレメントが存在している可能性が考えられた。これらの実験結果を確認するために、酵素分散法により得たウサギ大動脈平滑筋細胞の初代培養細胞で同様な実験を行ったところ、いずれの転写活性も他の非平滑筋細胞での転写活性よりも15倍以上の高値を示した。また、-92bpと-80bp間の欠失による転写活性の低下は非平滑筋細胞での低下に比べて顕著であった。
ガラクトシダーゼ・レポーター遺伝子を細胞に導入し、転写効率を補正した。まず、ウサギ大動脈平滑筋由来の血管平滑筋細胞C2/2にSM1/2が発現していることをRNase Protection Assayにて確認した後、転写活性を比較検討した。3.4kbp、1226bpのプロモーター領域を含むプラスミドの転写活性は、プロモーター・エンハンサーを含まないレポーター遺伝子、PGVB、に比較して50倍近い活性の上昇を認めた。-188bpまで欠失させたコンストラクトでも転写活性はそれほど変化しなかったが、より近位、特に、-92bpと-80bpの欠失で転写活性は顕著に低下し、この領域に活性増加作用領域が存在する可能性が示された。-72bpまで欠失させたプラスミドの転写活性はPGVBの9倍程度あるので、より近位に必須のシスエレメントが存在している可能性が考えられた。これらの実験結果を確認するために、酵素分散法により得たウサギ大動脈平滑筋細胞の初代培養細胞で同様な実験を行ったところ、いずれの転写活性も他の非平滑筋細胞での転写活性よりも15倍以上の高値を示した。また、-92bpと-80bp間の欠失による転写活性の低下は非平滑筋細胞での低下に比べて顕著であった。