| 内容要旨 | | 赤痢は細菌性下痢症の1つであり、発展途上国においては現在でもなお下痢が乳幼児の主要な死亡原因であり、細菌性赤痢は依然として根絶すべき対象となっている。赤痢は時として大流行を引き起こすこともあるが、その原因として多剤耐性菌の増加が治療を困難にさせており、今なお十分な効果を期待できるワクチンがないことも挙げられる。赤痢菌の感染を防御し、有効な予防法を確立するためには赤痢菌の病原性の解明が重要であると考えられる。赤痢菌は経口的に飲食物を介して感染し、腸管下部に到達した赤痢菌は腸上皮細胞に付着した後、非食細胞である腸上皮細胞へ貪食能を誘発し細胞質内へと侵入する。侵入した菌は、腸上皮細胞内で増殖し活発に運動しながら隣接細胞へ伝播拡散していき、やがて腸上皮細胞は炎症、剥離を生じ粘血性の下痢を引き起こす。この細胞侵入と拡散は赤痢菌の主要な病原性であると考えられ、前者を1次細胞侵入性、後者を2次細胞侵入性と本研究では呼ぶことにする。 この1次及び2次細胞侵入性に必要な遺伝情報の大部分は、本菌の保有する230-kbの巨大プラスミド上に存在する。現在までにこのプラスミド上にビルレンスに関わる様々な遺伝領域が同定されている。virB遺伝子、領域2、icsB遺伝子、領域3、領域4、領域5はクラスターとして存在している。この中で領域2-5は1次細胞侵入性に関与する遺伝領域と考えられており、領域2にはIpaB,IpaC,IpaDと呼ばれる3つの抗原蛋白質がコードされている。また領域3,4,5はIpa蛋白質の分泌に関与していると考えられている。このクラスターとは離れた位置に2次細胞侵入性に関与するvirG,virK遺伝子、またこれらの遺伝子の発現を調節するvirF,virB遺伝子などが存在する。1次細胞侵入性に関与する領域2-5は約31kbというDNA断片上にクラスターとして存在するため、その分子遺伝学的な解析は困難であった。そこで、本研究では抗原蛋白質IpaB,IpaC,IpaDをコードする領域2とIpaB,IpaC,IpaD蛋白質の分泌に関与していると考えられている領域5の分子遺伝学的解析を試み、領域2と領域5の赤痢菌の1次細胞侵入における役割について考察した。 上皮細胞侵入時に最も直接的な役割を果たしていると考えられているIpaB-Dをコードする領域2の遺伝子群の構造と機能を明かにするため、まずミニセル法による遺伝子産物の解析と領域2を含む7663bpの全塩基配列を行なった。その結果、領域2には24,18,62(IpaB),41(IpaC),37(IpaD)kDの5つの蛋白がこの順番にコードされていることが明かになった。また領域2のビルレンス回復試験と転写生成物の解析により、領域2は複合2オペロンとして存在することが示唆された。これら2つのオペロンのうち1つは24kD,18kD蛋白質,IpaB,IpaCをコードする遺伝子から成り、もう1つのオペロンはipaD遺伝子をコードし、さらにこれら2つのオペロンを含む領域2全体も1つのオペロンを形成していることが示された。また、各遺伝子にそれぞれフレイムシフト変異を導入しビルレンスとの関連性を調べたところ、ipaB,ipaC,ipaD遺伝子は細胞侵入性に関与しており、24kD,18kD蛋白質をコードする遺伝子は直接関与していないことが示唆された。そこで、IpaB,IpaC,IpaD蛋白質のアミノ酸の疎水性パターンを解析したところ、いずれも親水性の蛋白質でN末端に内膜通過に必要なシグナル配列もなく、また大腸菌のヘモリジン蛋白質の分泌に特有なC末端の膜貫通性領域も存在しなかった。さらにIpaB,IpaC,IpaD蛋白質の菌体局在を調べたところ、これらの蛋白質はいずれも外膜にはほとんど存在せず大部分が細胞質とペリプラスムに存在していた。しかし、これらの蛋白質は抗原蛋白質であり、現在までに上皮細胞侵入時に赤痢菌の菌体表面または菌体外に分泌され、その分泌が赤痢菌の細胞侵入性には不可欠であることが明かになっている。従って、これらの分泌には他の蛋白質が介在していることが強く示唆された。事実、領域2の上流21kbの領域3,4,5に変異を有する赤痢菌はIpa蛋白質の菌体内発現には全く異常がないにも関わらず、Ipa蛋白質の培養上清への分泌が抑制され、細胞侵入性が失われていた。そこで、領域3,4,5のうち領域5の遺伝子解析を行ない、Ipa蛋白質の分泌にどのように関与しているかを調べた。 領域5のビルレンスに必要な全遺伝領域をクローニングし、塩基配列を決定したところ、領域5には11個のオープンリーデイングフレーム(ORF)が検出された。推定アミノ酸から算出される分子量はそれぞれ15.1,47.5,13.2,33.0,33.4,24.2,9.4,28.5,39.9と10.4kDであった。またビルレンス回復試験の結果、領域5は1つのオペロンから成り、ORF-2からORF-9までの8個の遺伝子が細胞侵入性に必須であることが明かになった。さらに各IpaB,IpaC,IpaD特異抗体によるコロニーイムノブロットにより、8個のORFのうち7個(ORF-2とORF-5から-9)はIpaB,IpaC,IpaD蛋白質の分泌に、残りのORF-4はIpaBとIpaDの分泌に必要であることを明らかにした。そこでORF-2から-9の遺伝子をそれぞれspa47(surface presentation of plasmid antigen),spa13,spa32,spa33,spa24,spa9,spa29,spa40と命名した。Spa蛋白質のアミノ酸は、様々な病原性細菌あるいは非病原菌の蛋白質との間にホモロジーを有することが明かになってきている。これらの蛋白質は赤痢菌のspa領域と同様クラスターとして存在しており、遺伝子の配列も類似している。また、これらの蛋白質も病原性あるいは菌体表面構造物に関する蛋白質の細胞内から細胞外への分泌に必要であると考えられているが、それぞれの菌において分泌される蛋白質には類似性は見られず、むしろ分泌される蛋白質は各々特異的であることが知られている。また、赤痢菌の領域3,4にもIpa蛋白質の分泌に関するmxiと呼ばれる遺伝領域が存在することが明かになっており、これら領域3,4にコードされる複数の蛋白質もSpa蛋白質と共に細菌の輸送蛋白質との間に部分的なホモロジーが認められる。従って、spa領域とmxi領域の一部は細菌間で高度に保存され、これらの領域全体はいずれも蛋白質の分泌機構に関与している点で共通している。 グラム陰性菌の蛋白質の分泌機構としては従来、大腸菌のヘモリジン蛋白質に代表されるsec非依存型の分泌機構(TypeI型)とsec依存型で分泌蛋白質のシグナル配列による分泌機構(TypeII型)の2つの系が知られている。しかし、spa領域とmxi領域を代表とする蛋白質の分泌系はそれら2つの既知の分泌機構とは全く異なる新しい第三の分泌機構であることが明かになりつつある。この新しい分泌機構による蛋白質の分泌メカニズムはまだ解明されておらず、本研究で得られたSpa蛋白質の遺伝情報とその機能解析の結果は今後新しい蛋白質分泌機構の解明の基礎として重要であると考えられる。 |
| 審査要旨 | | 本研究は、赤痢菌が保有する230kbプラスミド上に存在する、細胞侵入性遺伝子群のうち、ヒト腸管上皮細胞への侵入に必要な遺伝領域(ipa領域及びspa領域)の分子遺伝学的解析を行ったものである。 その結果、ipa領域から産生される、IpaB,C,Dという3つの抗原性蛋白質はいずれもビルレンスに関与していることが明らかになった。また、これらIpa蛋白質は菌体外へと分泌され、その分泌が細胞侵入性には不可欠であった。しかし、これらIpa蛋白質はいずれも親水性蛋白質で、内膜通過に必要なシグナル配列も有していなかった。そこで、Ipa蛋白質の分泌に関与していると考えられるspa領域の遺伝子解析を行った。その結果、spa領域にコードされているspa47,spa13,spa32,spa33,spa24,spa9,spa29,spa40の8個の遺伝子はビルレンスに関与しており、これらの遺伝子はいずれもIpa蛋白質の菌体内から菌体外への分泌に関与していることが明らかになった。またその後、Spa蛋白質とホモロジーを持つ蛋白質が様々な動物性、植物性細菌に存在することが明らかになった。これらの蛋白質はSpa蛋白質と同様にクラスターとして存在しており、その遺伝子の配列も類似している。また、これらの蛋白質はいずれも病原性あるいは菌体表面構造物に関する蛋白質の菌体内から菌体外への分泌に必要であると考えられているが、分泌される蛋白質はそれぞれの菌において異なっている。従来、グラム陰性菌の蛋白質の分泌機構としては、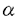 -ヘモリジンに代表されるsec非依存型(TypeI型)と分泌蛋白質のシグナル配列によるsec依存型(TypeII型)の2つの系が知られていた。しかしSpa蛋白質をプロトタイプとする蛋白質分泌機構はこれら従来の分泌機構とは異なる新しいIII型の分泌機構であることが明らかになりつつある。 -ヘモリジンに代表されるsec非依存型(TypeI型)と分泌蛋白質のシグナル配列によるsec依存型(TypeII型)の2つの系が知られていた。しかしSpa蛋白質をプロトタイプとする蛋白質分泌機構はこれら従来の分泌機構とは異なる新しいIII型の分泌機構であることが明らかになりつつある。 以上本研究では、従来行われていなかった赤痢菌の細胞侵入性に不可欠なIpa蛋白質をコードするipa領域と、Ipa蛋白質の分泌に関与しているspa領域の遺伝子の構造を明らかにした。この遺伝子解析は、赤痢菌の細胞侵入性を解明する上で最も基礎的なことであり、今後の研究を行うためにも重要であると考えられる。また、Spa蛋白質をプロトタイプとする新しい第III型の蛋白質分泌機構解明のためにも、本研究はその基礎として重要であり、学位の授与に値するものと考えられる。 |