学位論文要旨
| No | 212866 | |
| 著者(漢字) | 畝山,智香子 | |
| 著者(英字) | ||
| 著者(カナ) | ウネヤマ,チカコ | |
| 標題(和) | プリン受容体による巨核球の細胞質カルシウムオシレーシヨン | |
| 標題(洋) | ||
| 報告番号 | 212866 | |
| 報告番号 | 乙12866 | |
| 学位授与日 | 1996.05.08 | |
| 学位種別 | 論文博士 | |
| 学位種類 | 博士(薬学) | |
| 学位記番号 | 第12866号 | |
| 研究科 | ||
| 専攻 | ||
| 論文審査委員 | ||
| 内容要旨 | 巨核球は骨髄に存在する血小板の前駆細胞で、多核の大型細胞である。血小板は巨核球の細胞質がちぎれて生成するとされ、巨核球一つから血小板約4000個が生じるとされる。血小板には核はなく、従って血小板に含まれる成分はほとんどが巨核球由来であると考えられる。実際巨核球と血小板には糖鎖組成などの生化学的な類似点が多く報告されている。血小板は循環系の生理にとって極めて重要な成分であり、凝集する際に放出される生理活性物質はヌクレオチドから成長因子まで極めて多様である。しかし非常に小さくて凝集し易いため扱いの難しいものでもあり、単一細胞レベルでの研究は少ない。そのため血小板の代わりに巨核球を使った研究も行われている。本研究では単一細胞レベルでの研究を目的として、巨核球を材料に選んだ。 細胞の生理作用にとって細胞内Ca2+濃度の変化が極めて重要な意味を持つことは既に様々な系で明らかにされている。近年の技術の進歩により単一細胞レベルでの変動を解析することができるようになって、この「細胞内Ca2+濃度の変化」が個々の細胞では実は振動性の変化(オシレーション)である場合が多いことが知られるようになった。オシレーションの場合、刺激の強さは頻度に換算されるため、活性化に一定のCa2+濃度が必要なeffectorにとっては単純に刺激に応じてCa2+濃度が上昇する系よりも効果的にかつ繊細に刺激に応答することができると考えられる。こうした発見をもたらした新規技術の一つがpatch-clamp法である。従来のwhole-cell patch-clamp法をさらに改良して細胞内環境を維持したまま単一細胞の電気的応答を記録できるようにしたのがnystatin perforated法で、この方法によりpatch-clamp法の細胞生理学分野への応用が一段と広がることになった。さらに細胞外液を瞬時に交換できるY-tube法を採用することにより、早い反応の正確な記録を可能にした。本研究ではこのnystatin-perforated法を用いて巨核球のCa2+振動の性質及びその機序、生理的意義について検討した。 ラット骨髄から急性単離した巨核球のATP応答をnystatin perforated whole-cell patch clamp法により電圧固定下で記録すると振動性の電流が得られる。このATP誘発性の振動性電流は5mMのK+を含も標準細胞外液中では-80mVが平衡電位である。そこで外液+濃度と平衡電位との関係をプロットすると、Nernstの方程式から計算されるK+の平衡電位とほぼ等しく、この電流がK+により運ばれるものであることがわかる。さらにこのK+電流が細胞内Ca2+に依存することを示した。Conventional whole-cell法でシールを良くするために用いられることの多いEGTAを用いるとオシレーションが一過性の形になり、tailingも生じる。そのうえアゴニストに対する反応性もEGTA濃度に相関して減少する。このようなCa2+chelatorによるCa2+変動の緩衝作用は、光学的に単一細胞のCa2+濃度を測定する際に用いられるCa2+結合性の蛍光色素fura-2でもみられ、画像解析によるCa2+濃度測定で巨核球のCa2+オシレーションをとらえることはできない。EGTAを入れない場合には最初のうちはきれいな記録が取れるものの、時間とともに反応が鈍くなる(run-down)。こちらは細胞内分子がピペットの中に拡散していったために起こると考えられる。またK+チャンネルブロッカーによる阻害活性からこの電流がCa2+依存性K+チャンネルを通るものであることを明らかにした。以上の結果から巨核球のATP誘発性K+電流オシレーションは、非興奮性細胞のアゴニスト誘発性細胞内Ca2+オシレーションの一例であり、その記録にはここで用いたnystatin-perforated whole-cell patch-clamp法が現在のところ最も適していると結論できる。 次に用量相関について調べたところアゴニスト濃度に依存してもっとも大きく変化するのは頻度で、最大電流値はそれほど大きな変化を見せない。これらの性質は肝細胞での報告とほぼ一致する。またATPとADPとを比較したところ、頻度を指標にした場合、ADPはATPの約30倍の作用強度を示した。さらにその他のプリン受容体アゴニストについて調べた結果を表1に示す。ATPの非加水分解性類似体であるATP-   M)の場合[max frequency;0.21±0.02sec-1(n=7),max amplltude;637.5±68.8pA(n=8)]を1として表示した。各値は4-8検体のmean±S.E.M。nは細胞数 M)の場合[max frequency;0.21±0.02sec-1(n=7),max amplltude;637.5±68.8pA(n=8)]を1として表示した。各値は4-8検体のmean±S.E.M。nは細胞数次にこれまでに示したような巨核球のプリン受容体の血小板との違いの生理的意義について検討を加えた。ADPは血小板の弱いアゴニストとして凝集を促進することが知られている。血小板の強いアゴニストであるthrombinは多少反応時間が異なるものの巨核球にATPやADPとほぼ同様のCa2+オシレーションを誘発する。そこで巨核球の形態変化を走査型電子顕微鏡で観察した。Thrombinは巨核球に対しても凝集性の大きな形態変化をもたらすが、ATPおよびADPには見かけの形態変化を誘発する作用はなかった。Thrombinは血小板の細胞内Ca2+濃度の上昇とcAMP濃度の下降をもたらすことが知られており、そこでcAMP依存性プロテインキナーゼ(PKA)の阻害剤であるH-8で処理したところthrombin同様の形態変化がみられた。これは巨核球の急性の形態変化にはCa2+よりcAMPのほうが重要であることを示唆する。現時点では巨核球のCa2+濃度変化が引き起こす生理作用については同定できず、今後さらに検討を重ねる必要がある。 これまで多数の非興奮性細胞の細胞内Ca2+オシレーションに関する報告があり、それらの性質に応じていくつかのメカニズムに関する仮説が提案されている。巨核球の場合についてメカニズムの検討を行った。まず細胞外からのCa2+の流入がなくてもある程度のオシレーションの持続がみられることから、興奮性細胞のようなCa2+チャンネルの関与はないといえる。次いで細胞内にIP3を投与すると濃度に依存したオシレーションがみられ、このIP3誘発性オシレーションはIP3受容体アンタゴニストであるheparinで阻害され、Ca2+キレート剤BAPTAでも阻害される。またGTP- 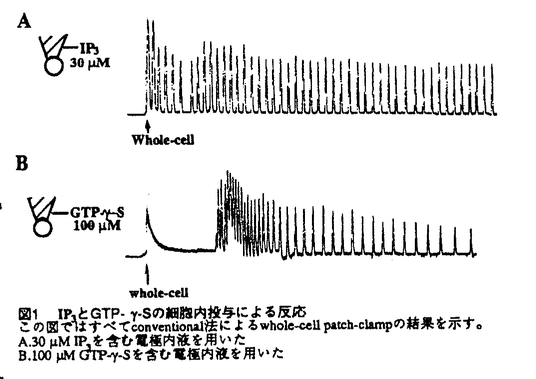  -Sの細胞内投与による反応M IP3を含む電極内液を用いた B.100M GTP--Sを含む電極内液を用いた -Sの細胞内投与による反応M IP3を含む電極内液を用いた B.100M GTP--Sを含む電極内液を用いたそこで振動を引き起こす要因の一つと考えられるprotein kinase C(PKC)の関与について検討した。PKCを活性化するphorbol myristate acetate(PMA)はATP応答を不可逆的に抑制するが、この抑制はPKCアンタゴニストstaurosporinで解除される。Staurosporinはオシレーションのパターンを一過性のつながった山の形に変える。これらのことからオシレーションのパターンにPKCが関与することが明らかである。さらにcAMPによる調節作用についても検討した。Adenylate cyclase活性化剤forskolinとphosphodiesterase阻害剤IBMXはいずれもATP誘発性K+電流オシレーションを抑制した。これらがIP3誘発性のオシレーションも抑制することから、その抑制点は細胞内Ca2+プールからのCa2+の放出以降にあると考えられる。 以上の結果についてまとめたのが図2である。このように巨核球のATP誘発性K+電流オシレーションはその再現性や定量性の良さから非興奮性細胞のCa2+オシレーションとしては極めて扱いやすいモデルとして、オシレーションのメカニズムや性質に関する様々な解析に有用である。 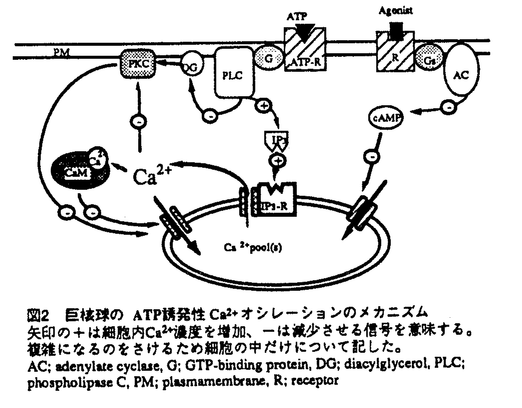 | |
| 審査要旨 | 細胞の生理作用にとって細胞内遊離カルシウムイオン濃度の上昇は極めて重要な意味を持つことが様々な系で明らかになってきている。その中でも近年の技術の向上により、単一細胞レベルで細胞内カルシウムイオン濃度が測定できるようになって、これまで単なる「濃度の上昇」としてとらえられていた現象の中には個々の細胞が振動性の変化(オシレーション)を示す場合があることが知られるようになってきた。こうした非興奮性細胞のカルシウムオシレーションは生体の機能に重要な役割を果たすと考えられるが、一見複雑な現象のため不明の部分が多い。本研究は改良された電気生理学の技術を巨核球に応用することで、カルシウムオシレーションがおこることを発見し、その性質や機構の解明を行った。 ラット骨髄から急性単離した巨核球のATP応答をnystatin-perforated whole-cell patch-clamp法で観察したところ、振動性の外向き電流を記録した。この電流はquinineなどのK+チャンネルブロッカーで抑制され、細胞内のCa2+をキレートすると抑制されることなどからCa2+-activated K+チャンネルを通るK+電流で、細胞内カルシウムイオン濃度の振動を反映するものであることを明らかにした。このK+電流振動はATP濃度に比例して頻度が増加するが、電流の大きさはATP濃度に関わらず比較的一定である。また巨核球のプリン受容体は、ATPよりADPが作用が強いなどこれまでに知られているどのサブタイプにも当てはまらず、新しいものであることを明らかにした。さらにその真のアゴニストはATP4-およびADP3-などの遊離の形のものであることや、P2Y受容体アンタゴニストが巨核球の受容体にもアンタゴニストとして働くことも見いだした。 巨核球のATP誘発性K+電流オシレーションをカルシウムオシレーションの指標として、そのメカニズムを解析した。巨核球の細胞内にconventional whole-cell patch-clamp法により直接IP3やGTP- 巨核球のATP誘発性K+電流オシレーションを修飾する因子について解析した。細胞内pHは酸性とアルカリ性ではそれぞれ別の作用点で促進と抑制の逆の作用を持つ。ミトコンドリア脱共役剤のFCCPは細胞の反応の程度によって異なる作用を示す。 また巨核球のATP誘発性K+電流オシレーションに対してadenineが増強作用を示すことを見いだし、adenineの生理作用について初めて報告した。 さらに巨核球の別のタイプのアゴニストであるthrombinとATPとの作用の違いについて比較することで、巨核球の形態変化に関与するセカンドメッセンジャーはCa2+ではなくcAMPであることを明らかにした。 以上のように、本研究では巨核球のカルシウムオシレーションという新たな現象を発見しそのメカニズムを解明するとともに、この実験系を利用して新規受容体を見いだし、細胞内カルシウム濃度調節機構の解明に役立てるなど、この系がカルシウムオシレーションのモデルとしても有用であることを示した。 従って本研究は薬理学、生化学の発展に寄与するところ大きく、博士(薬学)の学位にふさわしいものと判断した。 | |
| UTokyo Repositoryリンク | http://hdl.handle.net/2261/51001 |