緒言 Alzheimer病は痴呆を主症状とする神経変性疾患であり、病因はいまだ解明されていない。病理学的変化のうち細胞外の老人斑の蓄積が他の所見より早期に出現することから「老人斑の成因と神経変性のメカニズム」を解明することがAlzheimer病の病因に迫る有力なアプローチのひとつであると考えられている。
典型的な老人斑の中心部のコアは直径約10nmのamyloid線維からなる。老人斑の成因を探索するため、amyloidの構成成分が精力的に研究され、主構成成分としてA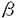 (タンパク、/A4)とその前駆体タンパクAPPが同定された。
(タンパク、/A4)とその前駆体タンパクAPPが同定された。
最近A以外のamyloidの構成成分タンパクとしてnon-A component of Alzheimer’s disease amyloid(NAC)が発見され、その前駆体NACPが同定された。NACはSDSによってamyloid線維から溶出しないことから、amyloidの基本構成成分と考えられ、この点で他の組織化学的に発見されたSDS可溶性の付随的amyloid成分とは異なる。しかし、NACとNACPに関する研究はいまだほとんどなされていない。
本研究では、NACとNACPのAlzheimer病病態への関与を調べることを目的に以下の項目について検討した。
1.NACPの脳内局在
2.Alzheimer病におけるNACPの変化
3.NACのamyloid形成能
4.NACPのAへの結合性
本論1.NACPの脳内局在 NACPの脳内局在を、NACPに対する特異的抗体を用いて検討した。
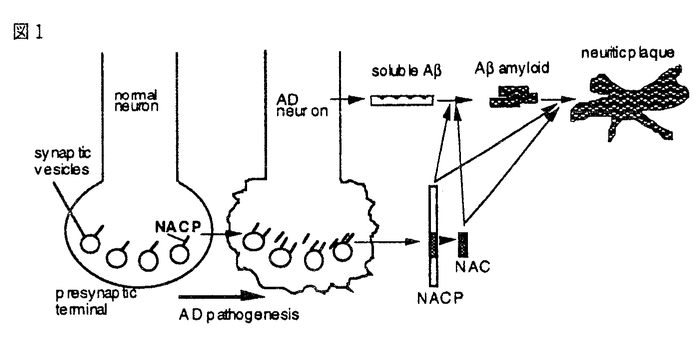
NACP C端とN端に対するポリクローナル抗体を作成した。両抗体ともラットおよびヒト脳の主に可溶性画分に19kDaのNACPを特異的に認識した。ラット脳切片を両抗体を用いて免疫組織化学的に検討したところ、嗅球、大脳皮質(II,III,およびV層)、線条体、側座核、海馬および小脳皮質が強く染色された。
光学顕微鏡下に観察すると、神経細胞のneuropilが点状に染色されていた。しかし、神経細胞の細胞体、glia細胞、および血管系細胞は染色されなかった。電子顕微鏡下に組織を観察すると、presynaptic terminalのsynaptic vesicleにNACP陽性の免疫反応が見られた。
Synaptic vesicle結合タンパクのsynaptophysinとNACPの共存状態を観察したところ、大脳皮質、海馬、基底核、嗅球、および視床では両タンパクの共存比率が高かった。小脳核と脳幹では、軸索樹状突起間synapseに両タンパクは共存していたが、軸索細胞体間synapseでは共存比率が低かった。
ラット脳各部位のNACPを定量した。NACPは嗅球、前頭大脳皮質、海馬、および線条体に多量に存在し、小脳、中脳、および延髄・橋では少量存在し、脳下垂体にはほとんど存在しなかった。この結果は、免疫染色におけるNACPの分布観察の結果と一致していた。
これらの結果より、NACPは中枢神経系の神経細胞presynaptic terminalのsynaptic vesicleに分布していること、Alzheimer病で病理的変化が高度な嗅球、大脳皮質、海馬といった発生的に新しい終脳領域に多量に発現していることを示した。Synapse特異的タンパクのNACPがamyloid構成成分の前駆体であることは、synapseの変性とamyloidの形成が強く関係していることを示唆する。
2.Alzheimer病におけるNACPの変化 Synaptic terminalの障害がAlzheimer病における痴呆程度とよく相関すること、またこの障害がAlzheimer病発症初期から観察されることから、Alzheimer病患者脳においてpresynapticタンパクであるNACPが変化していることが予想された。
Alzheimer病患者死後脳を、その病態の進行度により3群に分け、これに正常(control)脳を加え計4群とした。各脳のmidfrontal cortexの可溶性画分中のNACP免疫反応をNACP量として定量した。その結果、mild群のNACP量はcontrol群に比較して有意に増加していた。
Alzheimer病ではsynapse変性が進行し、synapse数が減少していることが知られている。ゆえに、NACPはAlzheimer病患者脳では減少していることが予想されたが、上の実験結果はその予想とは異なっていた。この点を検討するため、mid-frontal cortexのNACPの状態を免疫組織化学的に観察した。その結果、NACP陽性のsynapse数はAlzheimer病患者脳で有意に減少していたが、synaptic bouton1個あたりのNACP量を表すpixel densityは有意に増加していた。
これらの結果より、Alzheimer病を発症するとpresynaptic terminalにNACPが蓄積すること、そのためsynapse数は減少しても、synaptic boutonあたりのNACP量が増加するので、発症初期ではみかけのNACPの総量は増加することを示した。この知見は、Alzheimer病の病態にsynapse部分の変化が密接に関係していることを示唆する。
3.NACのamyloid形成能 NACのアミノ酸配列より、NAC自身にamyloidを形成する作用があることが予想されたので、35アミノ酸からなるNAC(NAC35)を人工的に合成し、そのamyloid形成能を検討した。
NAC35の濃度および温度依存的な重合が、濁度の上昇により示された。この溶液をSDS-PAGEで展開したところ、SDS不溶性の重合体の存在が観察された。
重合体をCongo redで染色し偏光下に観察すると、amyloid特有の複屈折が観察された。また電子顕微鏡下に観察すると直径約10nmの線維状の重合体が観察された。この線維形状はAlzheimer病のamyloidに酷似していた。
NACを特異的に検出するanti-NAC-X1でAlzheimer病患者脳切片を免疫染色すると、老人斑のamyloidが染色されたことから、amyloidにNACが存在することが組織学的に証明された。またこのNACはamyloidに一様に存在するのではなく、amyloid凝集体の中心部に高濃度で存在していた。
これらの結果より、NACはAと同様に自己重合してamyloid線維を形成すること、またNACはamyloidの中心部に存在することを示した。これらの知見は、NACがAlzheimer病amyloid形成の促進因子として働く可能性を示唆する。
4.NACPのAへの結合性 NACとamyloid主構成成分であるAの関係を検討するためにNACPのAへの結合性を検討した。
各種Aをニトロセルロース膜上に固定、NACP溶液中でインキュベートし、各Aに結合したNACPを定量した。NACPの結合量はA1-38>A25-35>A1-28であった。またNACPのsplice valiantであるNACP103-130を欠いたNACP112にも同様にA1-38との結合が観察された。NACP112からNAC部分を欠いたNACP ではA1-38との結合が消失した。
ではA1-38との結合が消失した。
両者の結合部分を特定するため、NACPとAとの結合に対するNACPおよびAのフラグメントの拮抗作用を検討した。溶液中にA25-35またはNACP81-95を加えるとA1-38とNACP112の結合は抑制されたが、他のフラグメントは抑制しなかった。
次にNACPがA aggregationを促進するかどうかを検討した。A1-38溶液にNACPを添加してインキュベートした後、混合溶液を遠心し、上清のA量をdotblotして定量したところ、上清中のA量はNACPを添加していないものに比較して有意に減少していた。沈殿物はthioflavin-Sで染色され、amyloid様の染色性を示した。
これらの結果より、NACPはAと結合するが、この結合にAではC端側25-35が、NACPにおいてはNAC部分のC端側NACP81-95が重要であることを示唆した。また、NACPのA溶液への添加によりamyloid形成が促進されることを明かにした。
 component of Alzheimer’s disease amyloid(NAC)の研究 : その脳内局在とアミロイド形成への関与
component of Alzheimer’s disease amyloid(NAC)の研究 : その脳内局在とアミロイド形成への関与