本研究はHIVを含むレンチウイルス、HTLV-1を含むオンコウイルスなど全てのレトロウイルスの複製過程において重要な役割を果たしている非スプライスRNA及びスプライスRNAの調節機構を解明することを目的としている。構造の単純なマウス白血病ウイルスを用いて以下の結果を得ている。 1.マウス白血病ウイルスにおいてgag遺伝子のみを持つような変異ウイルス(G3.6)では野生株ウイルスの約十分の一量のメッセージしか見られない。しかし、これにpol領域途中からEnv領域をつなぐと(GE6.4)メッセージレベルは野生株の発現量になる。この組換体でEnv領域に終止コドンをいれて蛋白をコードしないようにしても発現量は変わらない。しかし、スプライスアクセプター(SA)を欠失させたり(GEdSA)逆転させると(GEinvSA)メッセージ量はG3.6レベルに低下する。このG3.6にenv遺伝子のスプライスアクセプターを付加すると(GSA)mRNAのレベルは野生株とほぼ同じレベルまで回復する。このことからgagメッセージの発現のためにはSAが重要な役割を果たしていることが示唆される。 2.スプライスドナーは欠失させても(GEdSD)メッセージの発現は変わらず高くスプライスドナーはmRNAの発現レベルに影響しない事が示された。SAをG3.6のGag遺伝子領域の下流に挿入した際、SAを逆方向に挿入した場合(GinvSA)もメッセージ量の回復が見られた。即ち、逆向きでもmRNAメッセージ量を回復出来る事が示されたが、この場合にはaberrantにスプライスされたRNAが見られた。 GEタイプでSAを逆向きに挿入したGEinvSAとG3.6タイプでSAを逆向きに挿入したGinvSAを比較するとGEinvSAはGinvSAの全シークエンスを有し、発現の悪いGEinvSAには発現のよいGinvSAの全シークエンスが含まれている。したがって必要なのは特定のシークエンスではなくて特定の機能と考えられた。そこでメッセージの発現に必要なのはauthenticなSAのシークエンスなのではなくて機能しうるSAであると解釈される。また、温度感受性の実験から実際にスプライシングが起こる事は必要でなく機能しうるSAが存在することが重要であることが示された。 3.gag遺伝子領域をネオマイシン耐性遺伝子と交換した場合(neoEnv、neo-dE)はmRNAの発現レベルは高水準であり、このmRNAメッセージがSAに依存するという現象は一般的現象ではなくてgag遺伝子に特異的現象であることが示された。またgag遺伝子領域内に非スプライスRNAの発現量をSA依存にするシスエレメントが存在することが示された。 4.野生株や野生株に近いゲノムを持つウイルス( Wt:親株が野生株のRT領域に小欠損を導入したもの、fswt:親株が野生株のRT領域にフレームシフトを導入したもの)でSA領域を欠失(WtdSA,fswtdSA)させたり、逆転(WtinvSA,fswtinvSA)させてもコピーあたりのmRNAメッセージ量は著しく低下した。従ってゲノム長のプロウイルスにおいても非スプライスメッセージの発現にSAが重要な役割を果たしていることが確認された。 Wt:親株が野生株のRT領域に小欠損を導入したもの、fswt:親株が野生株のRT領域にフレームシフトを導入したもの)でSA領域を欠失(WtdSA,fswtdSA)させたり、逆転(WtinvSA,fswtinvSA)させてもコピーあたりのmRNAメッセージ量は著しく低下した。従ってゲノム長のプロウイルスにおいても非スプライスメッセージの発現にSAが重要な役割を果たしていることが確認された。 5.これら全てのプロウイルスは同じプロモーターにより転写されているのでRNAの発現の違いは転写後にもたらせると考えられる。SAの欠如によりmRNAの安定性が低下している可能性を考え、RNAの安定性を30度と37度で調べたがSAの有無によるRNAの安定性の相違は見られなかった。次に、SAの欠如が核から細胞質への輸送に障害を与えている可能性を考えて検討した。核と細胞質を分画しそれぞれのmRNAをノーザンブロットで比較した。wtdSAは低温、高温ともに核内のRNAメッセージ量は親株wtとほぼ等しく核内へのRNAメッセージの蓄積は見られなかった。一方、fswtdSAにおいては低温、高温ともに核内のRNAメッセージは親株fswtより著しく低下しており細胞質のRNA量に低下していた。すなわちfswtdSAでは核内におけるRNAの安定性が低下しており、WtdSAでは何らかの理由によりmRNAが核から細胞質への輸送の過程で壊されるのではないかと考えられた。 以上本論文はマウス白血病ウイルスにおいて非スプライスRNAの安定的発現のためには機能しうるスプライスアクセプターが必要であることを明らかにした。またこの現象はウイルスのgag遺伝子に特異的現象でありgag遺伝子領域にSA依存にさせる配列のあることが示唆された。本研究はレトロウイルスのmRNA発現の調節機構解明に重要な貢献をなすと考えられ学位の授与に値するものと考えられる。 | 
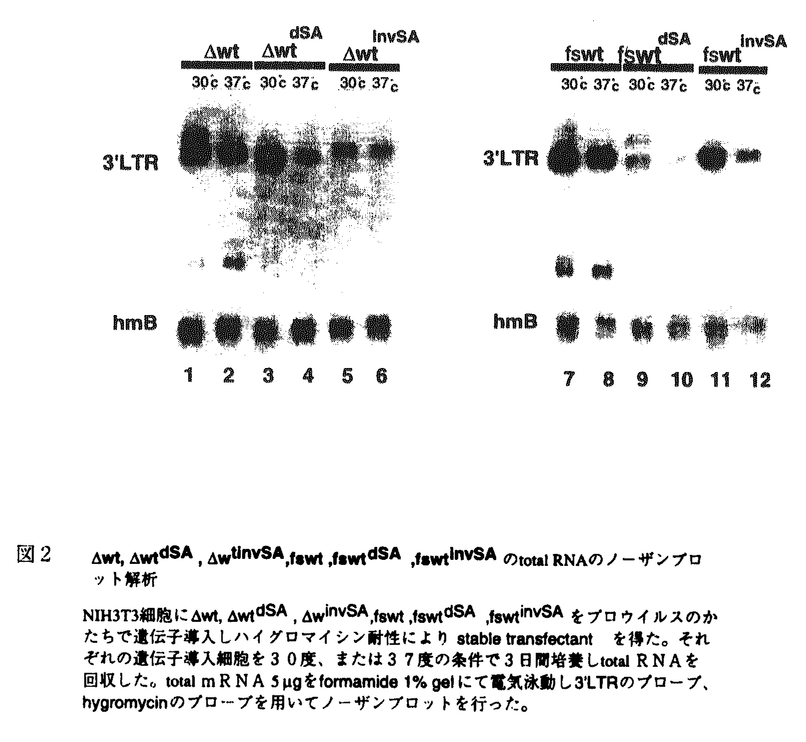
 gをformamide1%gelにて電気泳動し3’LTRのプローブ、hygromycinのプローブを 用いてノーザンブロットを行った。
gをformamide1%gelにて電気泳動し3’LTRのプローブ、hygromycinのプローブを 用いてノーザンブロットを行った。