アトピー性皮膚炎患者は、外来抗原(アレルゲン)に対して、IgE抗体を産生し、掻痒を伴う慢性の皮膚症状を呈する。アトピー性疾患(アトピー性皮膚炎、気管支喘息、アレルギー性鼻炎等)の家族歴を有することが多い。抗原特異的なIgE産生には、免疫応答を規定するHLA分子の関与が想定される。HLA分子上に提示された抗原ペプチドを、CD4陽性ヘルパーT細胞が認識することにより、Th2細胞に分化し、IgE産生を促すと考えられる。アトピー性皮膚炎の発症においても、HLA分子による抗原提示の拘束性が想定される。また、アトピー性皮膚炎は、皮疹の広がりや重症度、治療への反応性、血中IgE値等から、必ずしも発症要因は均一でないと考えられる。このため、本研究では、血中IgE値が8,000U/ml以上の高値を呈し、臨床型も重症の患者を解析の対象とした。 抗原ペプチドがHLA分子上に提示されるまでの、細胞内での抗原処理機構(antigen processing)の解析が進み、この機構に関わる遺伝子もクローニングされてきている。この中で、TAP(transporter associated with antigen processing)遺伝子とHLA-DM遺伝子は、HLAクラスII遺伝子領域に遺伝子座が存在している(図1)。また、両遺伝子ともに、遺伝子多型を示し、抗原ペプチドの選択に関わる可能性が考えられる。アトピー性皮膚炎においても、アレルゲンと想定されるダニ抗原等の細胞内抗原処理機構の解明は、疾患発症機序の解明にも繋がると期待される。本研究では、HLAクラスII遺伝子(DRB1、DRB3、DRB4、DRB5、DQA1、DQB1、DPB1)、および、HLAクラスI分子拘束性の抗原処理機構に関わるTAP遺伝子、HLAクラスII分子拘束性の抗原処理機構に関わるHLA-DM遺伝子をとりあげ、各遺伝子多型の解析を行ない、アトピー性皮膚炎発症への、これらの遺伝子の関与を検討した。 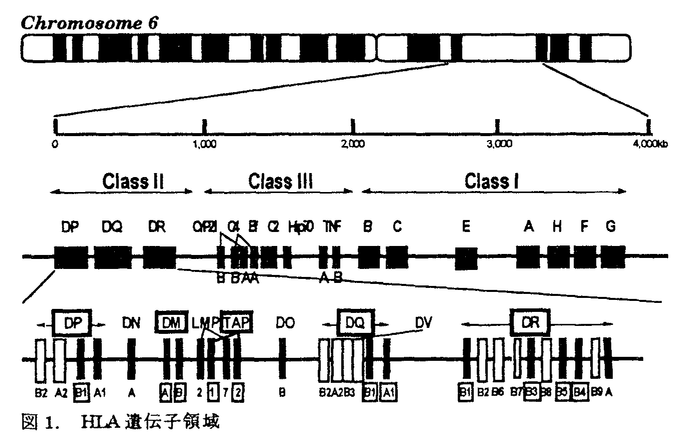 図1.HLA遺伝子領域 図1.HLA遺伝子領域 HLAクラスII遺伝子の解析では、患者群においては、DRB1*1302-DQB1*0604の増加を認めた(表1)。日本人においては、DR13は、強い連鎖不平衡によるA33-Cwblank-B44-DR13-DQ6のハプロタイプを形成しており、本研究での患者10名および対照3名の計13名とも、このHLAハプロタイプを認めた。さらに、アトピー性皮膚炎とDRB1*1302-DQB1*0604との相関をHLAクラスII分子のアミノ酸配列から検討した。HLA-DR分子上では、抗原ペプチドの側鎖を収容する5個のポケットが形成されている(図2)。DR分子上で、DRB1*1302と共通のアミノ酸配列を示すポケットを患者が持つ頻度は、第1ポケットでは、100%であり、このポケットでは 鎖86番目アミノ酸Glyがアミノ酸側鎖の結合を規定していると考えられる。第4ポケットおよび第7ポケットでは、Glu71、第9ポケットでは、Asp57、第6ポケットでは、Ser11、Ser13がペプチド結合に影響を与えていると推定される。DR-DQ間には、強い連鎖不平衡が存在し、DRB1*1302-DQB1*0604というハプロタイプは、例外を認めないため、現時点では、DR分子、DQ分子のどちらが第一義的に関与しているかは断定できない。アトピー性皮膚炎の発症に関わる想定されるアレルゲンの中で、ダニ抗原の解析が進んでおり、ダニの持つシステインプロテアーゼが、ダニ抗原の本体と考えられるに至っている。これら由来のペプチドが、おそらく皮膚ランゲルハンス細胞上に発現しているHLA-DR分子上の5個のペプチド結合ポケットを形成するアミノ酸の組合せによりに、HLA-DR分子への結合が規定されていると想定される。 鎖86番目アミノ酸Glyがアミノ酸側鎖の結合を規定していると考えられる。第4ポケットおよび第7ポケットでは、Glu71、第9ポケットでは、Asp57、第6ポケットでは、Ser11、Ser13がペプチド結合に影響を与えていると推定される。DR-DQ間には、強い連鎖不平衡が存在し、DRB1*1302-DQB1*0604というハプロタイプは、例外を認めないため、現時点では、DR分子、DQ分子のどちらが第一義的に関与しているかは断定できない。アトピー性皮膚炎の発症に関わる想定されるアレルゲンの中で、ダニ抗原の解析が進んでおり、ダニの持つシステインプロテアーゼが、ダニ抗原の本体と考えられるに至っている。これら由来のペプチドが、おそらく皮膚ランゲルハンス細胞上に発現しているHLA-DR分子上の5個のペプチド結合ポケットを形成するアミノ酸の組合せによりに、HLA-DR分子への結合が規定されていると想定される。 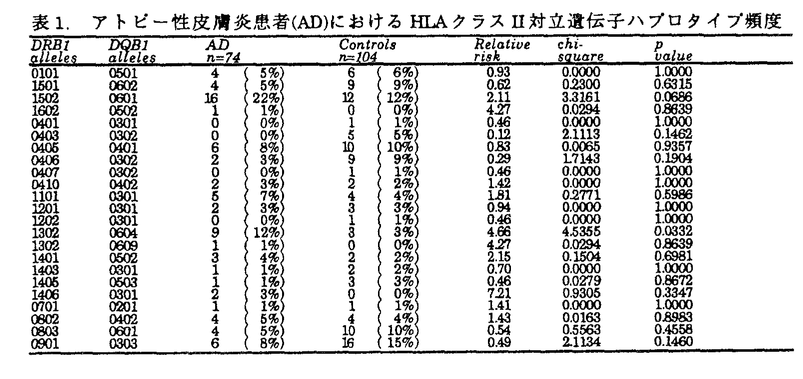 表1.アトビー性皮膚炎患者(AD)におけるHLAクラスII対立遺伝子ハプロタイプ頻度 表1.アトビー性皮膚炎患者(AD)におけるHLAクラスII対立遺伝子ハプロタイプ頻度 図2.HLA-DR分子上でのアトピー性皮膚炎の疾患感受性決定部位 図2.HLA-DR分子上でのアトピー性皮膚炎の疾患感受性決定部位 次に、抗原処理機構に関与するTAP遺伝子について解析を行なった。TAP分子は、HLAクラスI拘束性の抗原処理に際し、細胞質内抗原ペプチドを、粗面小胞体(ER)内に能動的に輸送する機能を持つ。TAP遺伝子に多型が存在し、TAP対立遺伝子の違いにより、輸送ペプチドの選択が、行なわれると想定される。既にHLAとの相関が判明している疾患で解析が、続々と報告されたが、現時点までで、調べられた限りでは、TAP遺伝子が第一義的に関連する疾患は、見出されていない。本研究でのアトピー性皮膚炎患者においても、特定のTAP対立遺伝子との相関は認められなかった(表2)。また、TAP2遺伝子では、687番目コドンが、終止コドン(TAG)の対立遺伝子(TAP2A、C、D、E)と、Gln(CAG)の対立遺伝子(TAP2B、G、H)が存在し、前者は、後者よりも、アミノ酸で17個短いが、Gln(患者23%、対照35%、RR=0.56)、終止コドン(患者77%、対照65%、RR=1.78)の頻度に、患者群と対照群で有意差は、認めなかった。TAP対立遺伝子とペプチドの輸送選択性についても解析途上であり、機能を含めた解析が今後必要であると考えられる。 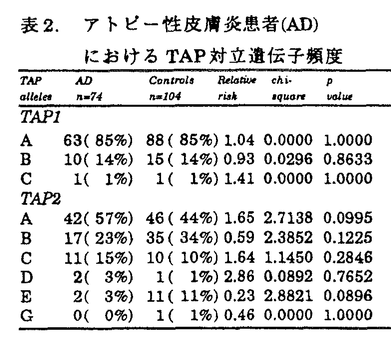 表2.アトピー性皮膚炎患者(AD)におけるTAP対立遺伝子頻度 表2.アトピー性皮膚炎患者(AD)におけるTAP対立遺伝子頻度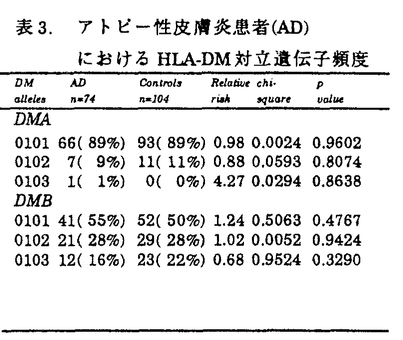 表3.アトピー性皮膚炎患者(AD)におけるHLA-DM対立遺伝子頻度 表3.アトピー性皮膚炎患者(AD)におけるHLA-DM対立遺伝子頻度 HLAクラスII分子拘束性の抗原提示に際して必要なHLA-DM分子の機能は、HLAクラスII拘束性の抗原提示に際して、CPL(compartment for peptide loading)内でHLAクラスII分子からインバリアント鎖(Ii鎖)を外し、抗原ペプチドが結合できる状態にすることと考えられる。HLA-DM対立遺伝子の違いにより、ペプチドのHLA分子の結合に違いがあると想定されている。現時点では疾患とHLA-DM対立遺伝子との相関の報告はなく、本研究で初めて解析が行なわれた。しかしながら、対立遺伝子頻度に、対照群と有意差は認められなかった(表3)。 HLAクラスII分子の抗原処理機構では、HLA-DM分子およびIi鎖の両者により、抗原ペプチドのHLAクラスII分子への結合が、支配されているため、今後、両者を統合した解析を進める必要があると考えられる。また、DMAおよびDMB対立遺伝子の違いが、Ii鎖あるいは抗原ペプチドの結合に与える影響については、未解決であり、分子の機能解析を含めて、明らかにする必要があると思われる。 本研究では、PCR-RFLP法を用いて、遺伝子多型の解析を行なった。PCR増幅産物中に制限酵素認識配列を作成するmismatch-PCR-RFLP法を開発し、PCR-RFLP法の欠点を補完することが可能となった。日本人で比較的出現頻度の高いDRB1*1501とDQB1*0604の対立遺伝子を同定するため、およびTAP遺伝子の解析に使用し、この過程で日本人で初めてDQB1*0609対立遺伝子を同定した。Mismatch-PCR-RFLP法は、今後、広く応用できる方法であると考えられる。 HLA分子への抗原ペプチドの結合様式の解析、結合しているペプチドのモチーフの解析が急速に進行している。HLA分子の側からみると、従来、MHC拘束性として認められてきた現象が、HLA分子上でペプチド結合可能なポケットを形成するアミノ酸配列に基づくことが明らかにされている。本研究でのアトピー性皮膚炎でも、DR分子、DQ分子上のポケットを形成するアミノ酸配列との相関が認められ、ダニ抗原等のアレルゲンの結合を拘束していると考えられる。抗原ペプチドの結合様式の解析、結合ペプチドのモチーフの解析等とともに、抗原処理および抗原提示機構の研究の進展を通じて、臨床医学においても、今後、疾患の発症機序、対応抗原の解明へと進むと期待できる。 |  鎖7-31-32-34-43-
鎖7-31-32-34-43-