膵は内胚葉性上皮由来の臓器であり、胎生期膵導管の前駆細胞から膵外分泌腺と膵内分泌腺が分化してゆくと考えられている。その分化の仕組み、分化を調節する因子等はまだ明らかではない。膵内分泌腺細胞がA(グルカゴン)細胞、B(インスリン)細胞、D(ソマトスタチン)細胞、PP(パンクレアティックポリペプチド)細胞へと分化してゆく仕組みもまだ明らかではないが、Hanahanらが一つのモデルを提唱している。そのモデルではグルカゴンを発現する細胞がまず出現し、複数の膵ホルモンを発現する細胞が混在する形で分化し、やがて単一のホルモンを産生する細胞に分化してゆくというものである。このモデルは概念的には理解しやすいモデルであるが、その後いくつかの合致しない点が指摘されており、膵内分泌腺細胞の分化のメカニズムに関しては混沌としているのが現状である。また分化を調節する因子に関してはほとんど解明されていない。 AR42J細胞は化学的に誘導されたラット膵腫瘍由来の細胞株で、膵外分泌腺細胞としての性格と神経内分泌腺細胞としての性格を合わせもっており、膵導管系上皮細胞に由来すると考えられている。デキサメサゾン処理により膵外分泌腺としての性格を強くすることは以前からよく知られていたが、TGF-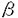 スーパーファミリーの一員であるアクチビンAをAR42J細胞に作用させると神経細胞に特徴的な細胞骨格を呈し、膵内分泌腺細胞に特徴的なチャネルを発現することを最近我々は明らかにした。アクチビンA処理したAR42J細胞は神経内分泌腺細胞としての性格を強くするのだが、膵内分泌腺細胞が膵ホルモンの分泌の他に様々な神経細胞様の性格(神経特異的マーカーの発現、細胞膜の電気的興奮性、B細胞をin vitroで培養するとニューロフィラメントを有する神経突起を伸長する)をもっていることを考え合わせると、AR42J細胞にさらに何らかの分化誘導因子を作用させれば、この細胞が膵内分泌腺細胞にまで分化する可能性が考えられる。この可能性を検証するために本研究を行った。 スーパーファミリーの一員であるアクチビンAをAR42J細胞に作用させると神経細胞に特徴的な細胞骨格を呈し、膵内分泌腺細胞に特徴的なチャネルを発現することを最近我々は明らかにした。アクチビンA処理したAR42J細胞は神経内分泌腺細胞としての性格を強くするのだが、膵内分泌腺細胞が膵ホルモンの分泌の他に様々な神経細胞様の性格(神経特異的マーカーの発現、細胞膜の電気的興奮性、B細胞をin vitroで培養するとニューロフィラメントを有する神経突起を伸長する)をもっていることを考え合わせると、AR42J細胞にさらに何らかの分化誘導因子を作用させれば、この細胞が膵内分泌腺細胞にまで分化する可能性が考えられる。この可能性を検証するために本研究を行った。 まずはじめにアクチビンA処理したAR42J細胞が膵ホルモンを産生しているかどうか、膵ホルモンのメッセンジャーRNA(mRNA)の発現をReverse transcription polymerase chain reaction(RT-PCR)法で検討した。インスリンやグルカゴンの発現はみられなかったが、アクチビンA処理によりPPのmRNAの発現を認めた。アクチビンA処理したAR42J細胞を抗PP抗体を用いて免疫細胞染色すると最大で約25%の細胞がPP陽性であったが、その数はアクチビンA処理72時間後が最大で、細胞が突起を伸長するにつれ減少した。またアクチビンA処理により、グルコーストランスポーター2(GLUT2)のmRNAの発現を認めた。アクチビンAはPP陽性細胞の出現を促すが、その作用は一過性であり、単独では膵内分泌細胞への弱い誘導作用しか持たないと考えられた。 次にアクチビンA以外にAR42J細胞を膵内分泌腺細胞に分化誘導する因子が存在するかどうかを抗インスリン抗体を用いた免疫細胞染色でスクリーニングした。Epidermal Growth Factor (EGF)、Transforming Growth Factor 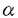 (TGF-)、Fibroblast growth factor(FGF),Insulin-like growth factors,ガストリン、コレシストキニン、Transforming Growth Factor (TGF-)、ベータセルリン等を検討したところ、EGFファミリーに属するベータセルリンがAR42J細胞の一部をインスリン陽性細胞に誘導した。このベータセルリンの効果は用量依存性で2nMで最大効果が得られ、約4%の細胞がインスリン陽性となった。ベータセルリンはまたAR42J細胞の一部をPP陽性細胞へと誘導した。RT-PCRによる検討では、ベータセルリンで処理したAR42J細胞はインスリン、PP、GLUT2、グルコキナーゼのmRNAの発現を認めた。グルカゴンの発現は認めなかった。インスリン陽性細胞はトルブタマイドに反応して細胞内カルシウム濃度が上昇した。これはATP感受性カリウム(KATP)チャネルの発現を意味する。ベータセルリンは増殖促進因子と考えられているが、AR42J細胞の増殖は促進しなかった。またAR42J細胞全体のアミラーゼ含有量には影響を及ぼさなかった。これはベータセルリンにより誘導されるインスリン陽性細胞、PP陽性細胞はごく一部の細胞であり、しかもこれらの細胞は増殖せず、大多数の房状に増殖する細胞にマスクされるためと考えられた。ベータセルリンの増殖促進作用はEGF受容体を介しての作用と考えられているが、AR42J細胞を分化誘導する作用はEGFやTGF-では再現されず、ベータセルリン固有の受容体を介しての作用の可能性が考えられた。しかし大部分のAR42J細胞はベータセルリンに反応せず、房状に増殖してアミラーゼを産生した。ベータセルリンに反応しない理由として、この細胞がポリクローナルな集団で、十分なベータセルリン受容体を発現している細胞がわずかしかない場合が考えられた。ベータセルリンの作用に加えて別の因子を作用させれば、さらに多くの細胞がベータセルリンに反応するようになる可能性がある。この観点から、AR42J細胞を神経内分泌細胞に分化誘導する作用のあるアクチビンAをベータセルリンと共にAR42J細胞に作用させ、その効果を検討した。 (TGF-)、Fibroblast growth factor(FGF),Insulin-like growth factors,ガストリン、コレシストキニン、Transforming Growth Factor (TGF-)、ベータセルリン等を検討したところ、EGFファミリーに属するベータセルリンがAR42J細胞の一部をインスリン陽性細胞に誘導した。このベータセルリンの効果は用量依存性で2nMで最大効果が得られ、約4%の細胞がインスリン陽性となった。ベータセルリンはまたAR42J細胞の一部をPP陽性細胞へと誘導した。RT-PCRによる検討では、ベータセルリンで処理したAR42J細胞はインスリン、PP、GLUT2、グルコキナーゼのmRNAの発現を認めた。グルカゴンの発現は認めなかった。インスリン陽性細胞はトルブタマイドに反応して細胞内カルシウム濃度が上昇した。これはATP感受性カリウム(KATP)チャネルの発現を意味する。ベータセルリンは増殖促進因子と考えられているが、AR42J細胞の増殖は促進しなかった。またAR42J細胞全体のアミラーゼ含有量には影響を及ぼさなかった。これはベータセルリンにより誘導されるインスリン陽性細胞、PP陽性細胞はごく一部の細胞であり、しかもこれらの細胞は増殖せず、大多数の房状に増殖する細胞にマスクされるためと考えられた。ベータセルリンの増殖促進作用はEGF受容体を介しての作用と考えられているが、AR42J細胞を分化誘導する作用はEGFやTGF-では再現されず、ベータセルリン固有の受容体を介しての作用の可能性が考えられた。しかし大部分のAR42J細胞はベータセルリンに反応せず、房状に増殖してアミラーゼを産生した。ベータセルリンに反応しない理由として、この細胞がポリクローナルな集団で、十分なベータセルリン受容体を発現している細胞がわずかしかない場合が考えられた。ベータセルリンの作用に加えて別の因子を作用させれば、さらに多くの細胞がベータセルリンに反応するようになる可能性がある。この観点から、AR42J細胞を神経内分泌細胞に分化誘導する作用のあるアクチビンAをベータセルリンと共にAR42J細胞に作用させ、その効果を検討した。 アクチビンAとベータセルリンを共にAR42J細胞に作用させると増殖はほぼ停止し、ほとんどの細胞が双極性の突起を伸長したが、中には丸いまま増殖しない細胞もみられた。約10%の細胞がインスリン陽性となったが、突起を伸長した細胞、円形の細胞の両者にインスリン陽性細胞はみられた。PP陽性細胞は円形の細胞にしかみられなかった。RT-PCRによる検討では、インスリン、PP、グルコキナーゼ、GLUT2のmRNAの発現を認めたが、グルカゴンの発現はやはり認めなかった。アクチビンAとベータセルリンを共に作用させても、全ての細胞がインスリン陽性となるわけではなかった。その原因としてAR42J細胞がポリクローナルな集団である可能性が考えられたため、クローンをスクリーニングすることにより、この中からアクチビンAとベータセルリンの作用によりほぼ100%インスリン陽性となるクローンAR42J-B20細胞を得た。この細胞を使ってインスリンが調節性の経路により分泌されるかどうかを検討した。高カリウム液、トルブタマイド、カルバコール、Glucagon-like peptide-1(GLP-1)の刺激によりインスリンが分泌され、正常B細胞にみられる調節性の分泌経路が保たれていることが示された。高濃度グルコースには反応しなかったが、これはアクチビンAとベータセルリンによって誘導されるGLUT2、あるいはグルコキナーゼの発現量が十分ではない可能性が考えられた。このAR42J細胞の系は我々の知る限りでは、アミラーゼ分泌細胞がインスリン分泌細胞へと分化する初めてのin vitroモデルシステムである。 AR42J細胞は膵腺房由来の腫瘍から化学的に誘導された腫瘍細胞ではあるが、この細胞を使って見られる現象は正常細胞で見られる現象と矛盾しない点が多い。例えば、AR42J細胞は分泌顆粒を有し、CCKなどの刺激でアミラーゼを分泌し、またインスリン、ガストリン、EGFなどで増殖が促進される。アクチビンAはデキサメサゾンで誘導されるAR42J細胞の外分泌細胞への分化を抑制するが、膵原基の組織培養ではアクチビンAは腺房の分枝形成を抑制する。AR42J-B20細胞は調節性のインスリン分泌経路を持っている。従ってこのAR42J細胞のモデル系を利用して得られる情報は正常細胞で見られる現象と矛盾しない可能性が高い。 本研究ではAR42J細胞を用いて以下の点を明らかにした。 1.ベータセルリンはAR42J細胞の一部をインスリン産生細胞に分化誘導し、その作用はアクチビンAにより増強された。ベータセルリンやアクチビンAは分化過程の膵内分泌前駆細胞や膵ラ氏島に発現しており、オートクリン・パラクリン的に作用し、膵内分泌腺細胞の形成や分化に生体内で関与している可能性が十分考えられる。ただしこれらの作用はin vivoではまだ検討されてはおらず、今後検討していかなければならない。 2.AR42J細胞はインスリン、PP陽性細胞にはなるが、グルカゴン陽性細胞にはならない。このモデル系ではPP陽性細胞を介してインスリン陽性細胞が出現すると考えられ、PPの発現がB細胞の分化に大きな役割を果たしている可能性が示唆された。 3.AR42J細胞はデキサメサゾンと共に培養すると外分泌線としての性格を強くし、ベータセルリン、アクチビンAと共に培養すると膵ホルモン産生細胞に分化する。膵導管上皮の前駆細胞と共通の性格を有しており、この細胞は膵の分化を検討できる初めてのin vitroモデルである。 本研究ではベータセルリンの分化誘導因子としての作用を初めて明らかにし、AR42J細胞が膵の分化を検討できる画期的なin vitroモデル系であることを示した。今後はこのモデル系を利用して、前駆細胞から内分泌腺細胞、外分泌線細胞への分化を決定する因子、膵内分泌前駆細胞からインスリン産生細胞へと分化する過程でそれを調節する因子等を探求してゆく予定である。 |